



Orkidéer er planter som tilhører orkidéfamilien. På norsk kalles også familien marihåndfamilien. Antall navngitte arter er om lag 29 000 og familien trolig den mest artsrike av plantefamiliene, avhengig av hvor mange arter man regner med i kurvplantefamilien. I Norge finnes omtrent 35 orkidéarter fordelt på 23 slekter, blant annet marisko, flueblom, nattfioler og flere typer marihånd.
Faktaboks
- Etymologi
- Navnet orkidé kommer av gresk ‘testikkel’, etter de to eggrunde rotknollene hos slekta vårmarihånd, Orchis. Navnet «marihåndfamilien» kommer fra formen på rotknollene hos slekta marihånd, som minner om hender.
- Også kjent som
- marihåndfamilien
- Vitenskapelig navn
- Orchidaceae
Orkideer er flerårige urter med varierende utseende og levevis. Det finnes orkideer i de fleste naturtyper på landjorda, men flest ulike arter finnes i tropiske strøk med høy temperatur og luftfuktighet.
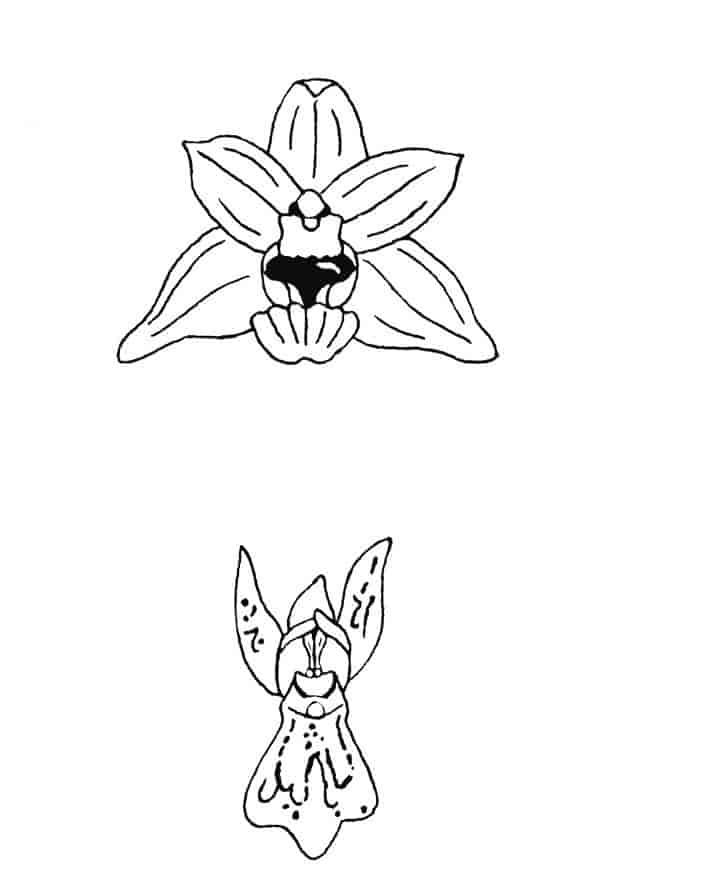
Blomstene hos orkideene er som regel fargerike og består av seks blomsterblad, ofte med ett av disse forma som en karakteristisk leppe.
Orkideer pollineres av dyr, hovedsakelig insekter, og noen av tilpasningene mellom blomst og pollinator er blant de underligste i naturen. Det finnes for eksempel blomster som etterligner insekters utseende og duft og blomster som etterligner andre blomster. Svært mange, kanskje halvparten, av orkidéartene narrer pollinatorene, idet blomstene ikke har noen belønning i form av for eksempel nektar.
Bruk av orkideer som prydplanter har økt mye de seinere årene og er nå en stor industri. Utover dette har orkideer samla sett liten økonomisk betydning, bortsett fra noen viktige unntak som vanilje og salep.















Kommentarer (2)
skrev Kjetil Skogen
svarte Kjell-Olav Hovde
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.