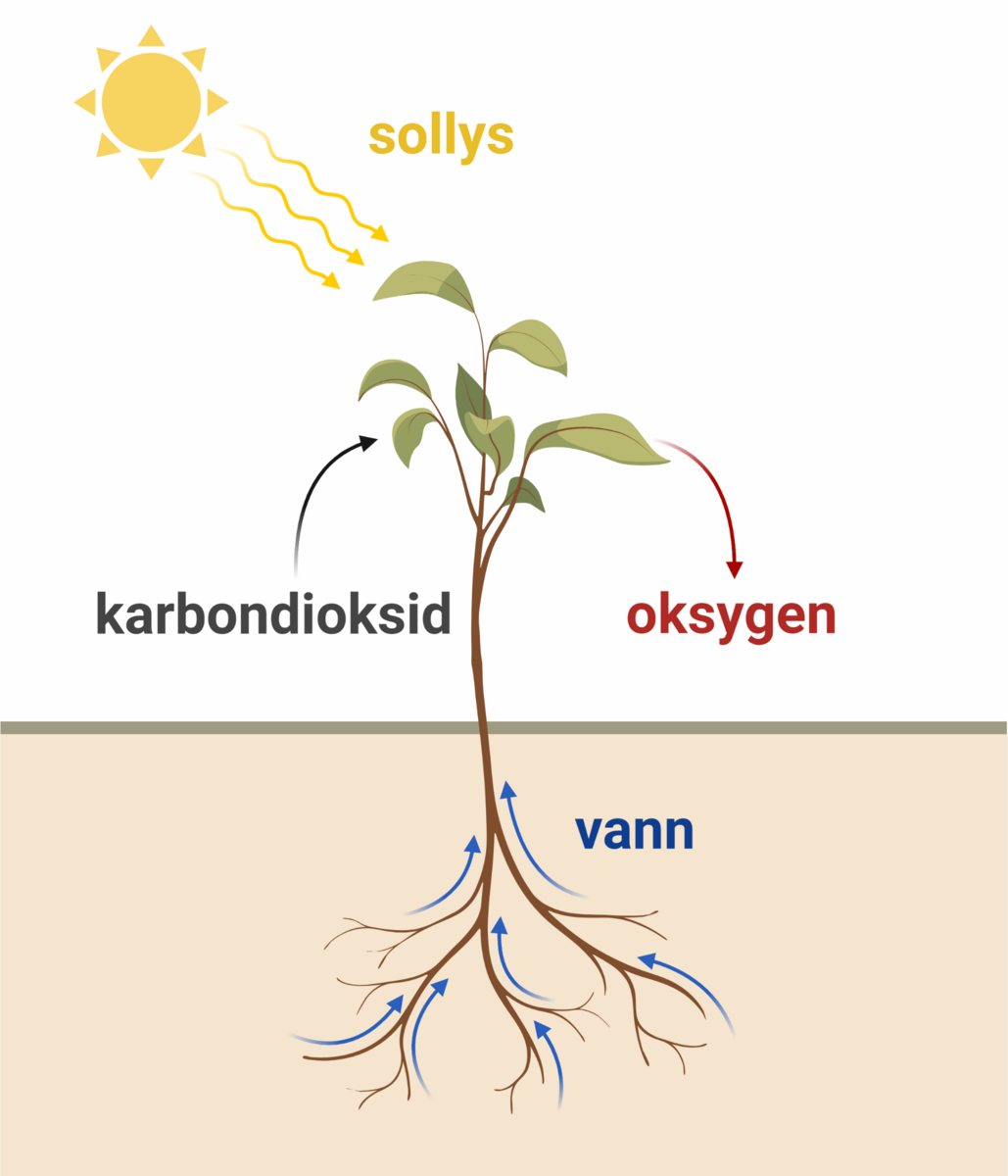
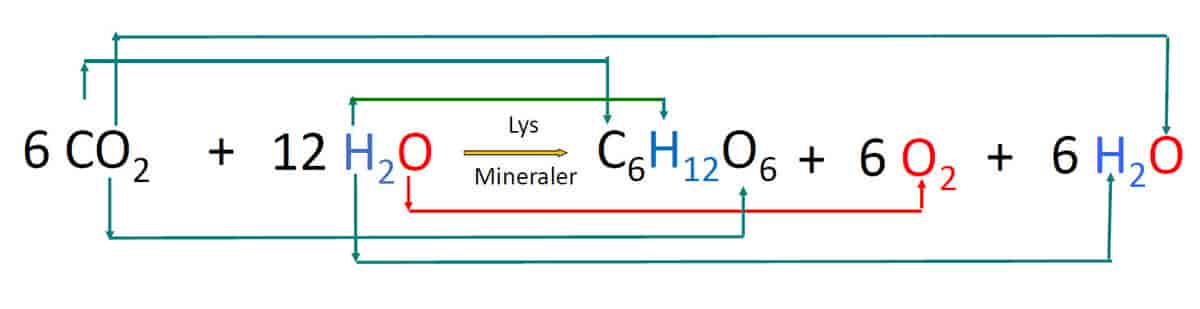
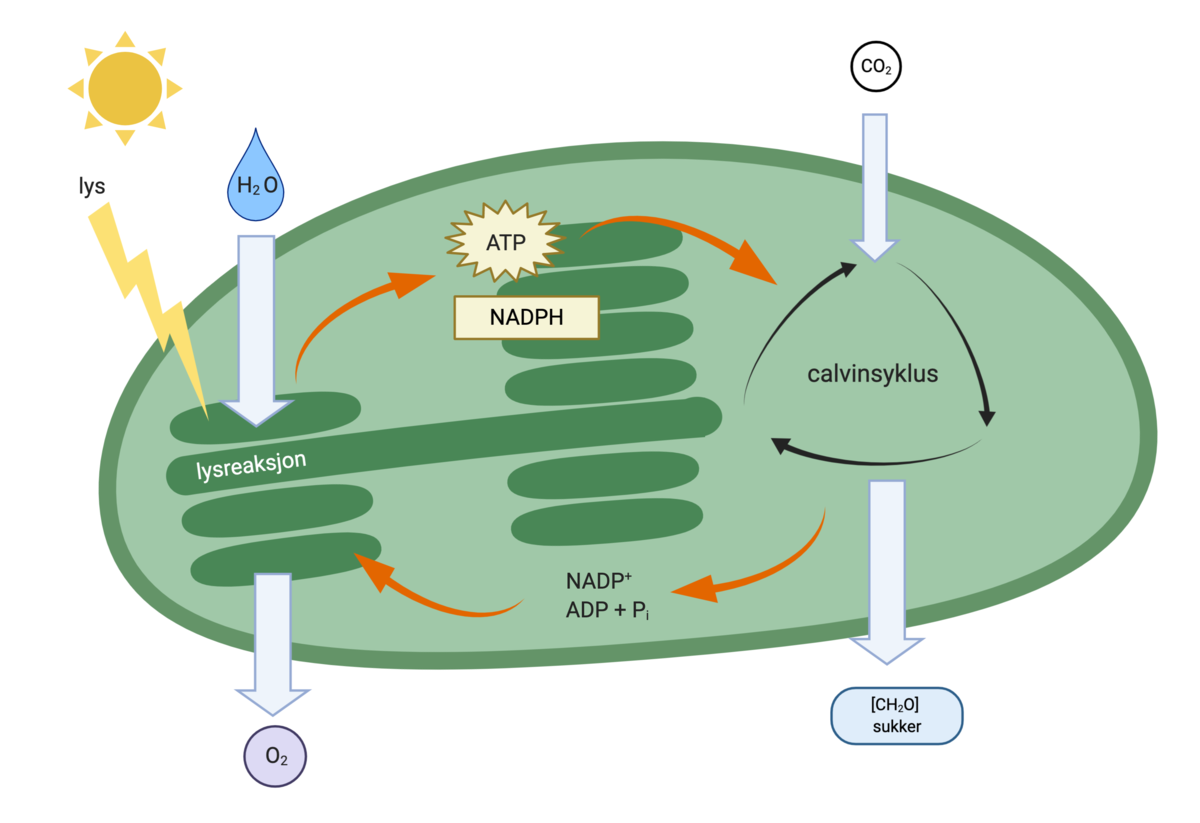
Fotosyntese er en kjemisk reaksjon. Kloroplastene kan sees på som plantecellenes laboratorium for å fange lysenergi. For å få dette til må plantene flytte elektroner og protoner, og spalte vannmolekyler. Inne i kloroplastene er det komplekse strukturer som sørger for at disse kjemiske reaksjonene skjer i riktig rekkefølge.
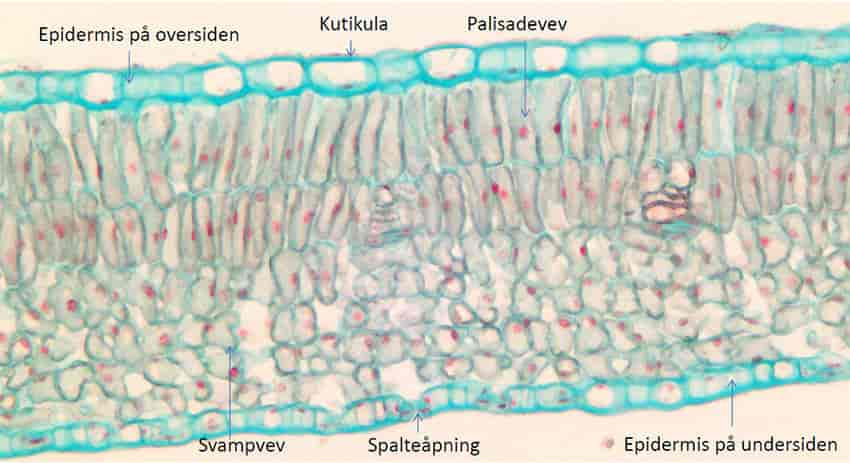
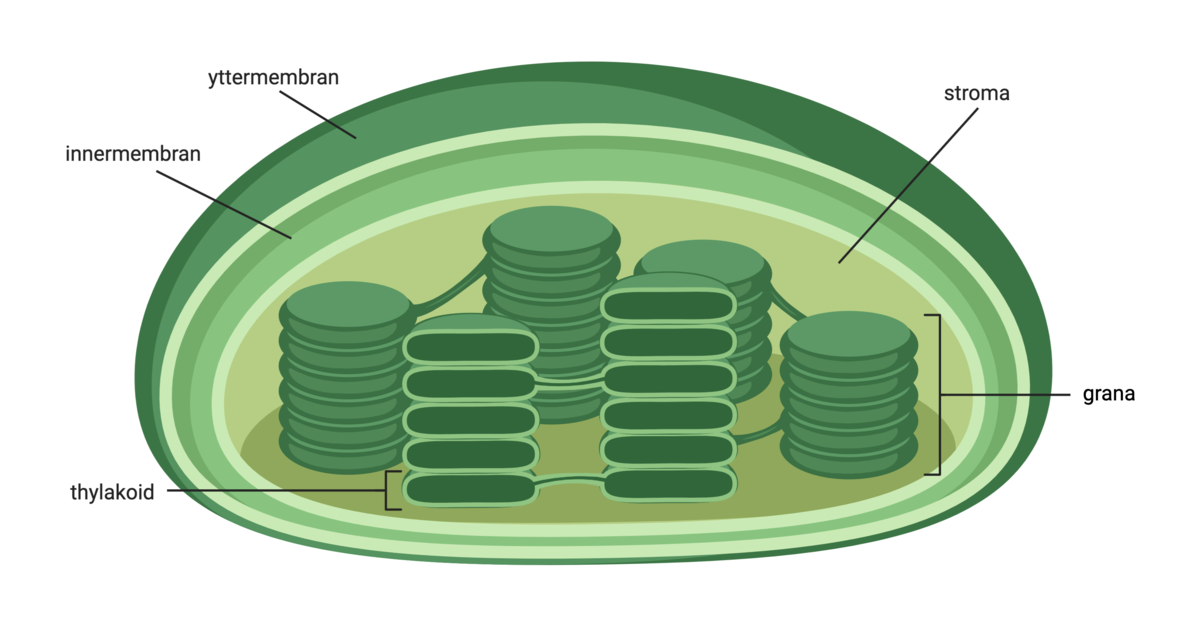
Kloroplastene inneholder noen skiver kalt tylakoider. Disse er delvis frittliggende og delvis formet som stabler. Tylakoidene er omgitt av en membran, og i denne membranen foregår viktige steg i fotosyntesen. Kun tylakoidmembranene har grønn farge, resten av kloroplasten er fargeløs og kalles stroma. Det grønne pigmentet klorofyll er plassert i tylakoidmembranen, sammen med andre fotosyntesepigmenter. Det er her energien fra lyset fanges.
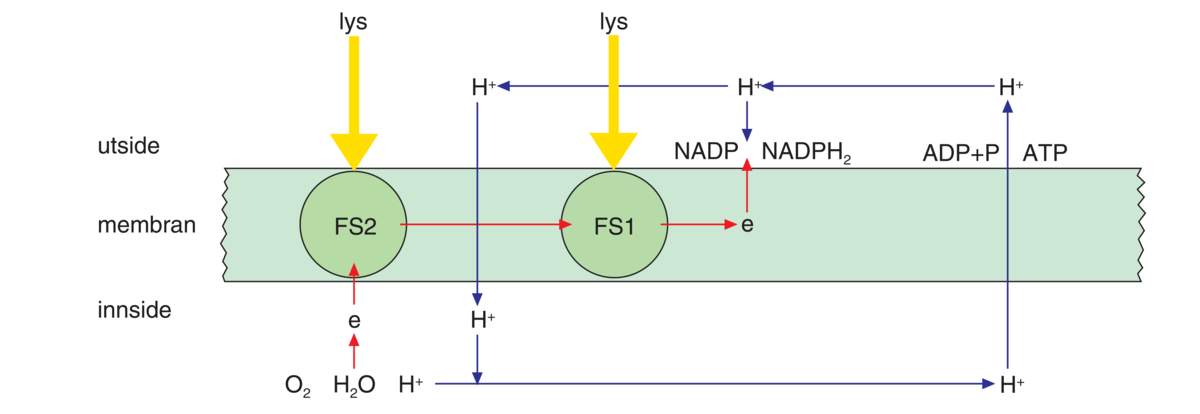
Fotosyntesepigmenter i tylakoidmembranene er satt sammen med proteiner i antennekomplekser. Det er disse antennekompleksene som fanger solenergi. Antennekompleksene er plassert rundt et reaksjonssenter med hundrevis av molekyler av klorofyll a. Energien fra sollyset som fanges opp av antennekompleksene blir sendt til reaksjonssenteret.
I reaksjonssenteret mottar klorofyll a energien fra sollyset, og et elektron fra magnesiumionet (Mg2+) i klorofyll a løsrives og sendes videre via en elektrontransportkjede. Klorofyll a finnes i to ulike varianter, og dette gir opphav til to ulike former for reaksjonssentre med noe ulik funksjon. Dette kalles fotosystem 1 og 2, og gir opphav til to litt forskjellige elektrontransportkjeder.
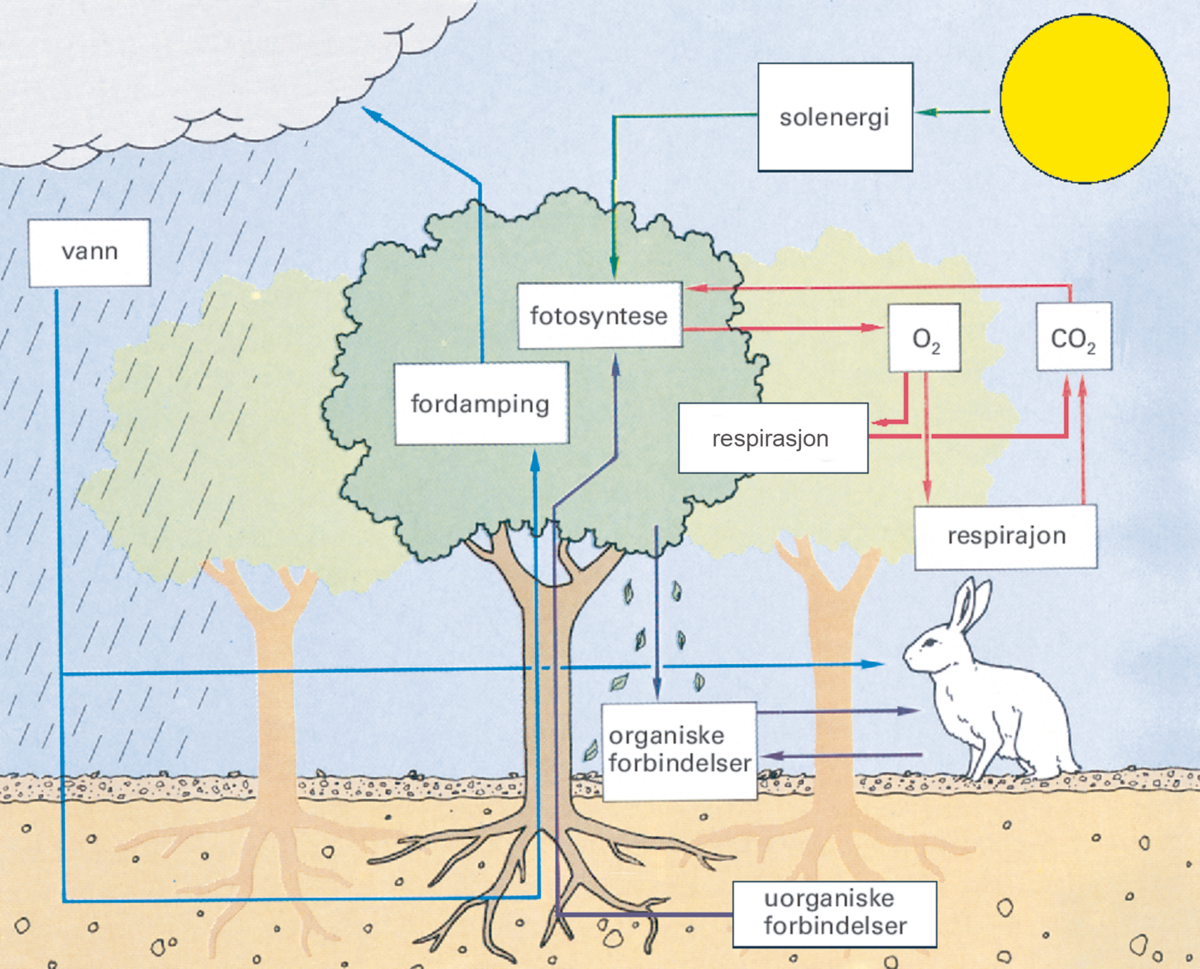
Planten trenger både fotosystem 1 og 2 for å gjennomføre alle prosessene i fotosyntesen. I elektrontransportkjedene fra fotosystem 1 og 2 dannes både ATP, som lagrer kjemisk energi, og NADPH, som kalles reduksjonskraft, og trengs til produksjonen av karbohydrater. I fotosystem 2 blir vannmolekyler spaltet, slik at oksygengass blir frigitt. Oksygenet som blir frigitt i fotosyntesen kommer fra vann, og ikke fra karbondioksid.









Kommentarer (6)
skrev Malin Pedersen
svarte Halvor Aarnes
svarte Malin Pedersen
skrev Petter Ulleland
Er ikke "respirasjon" feilstavet i bildet øverst til høyre?
svarte Marit M. Simonsen
Hei! Tusen takk for at du gjorde oss oppmerksom på dette, nå er det rettet. Hilsen Marit i redaksjonen
svarte Petter Ulleland
Det er fortsatt en stavefeil til stede ... det var to, nå er det en.
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.