
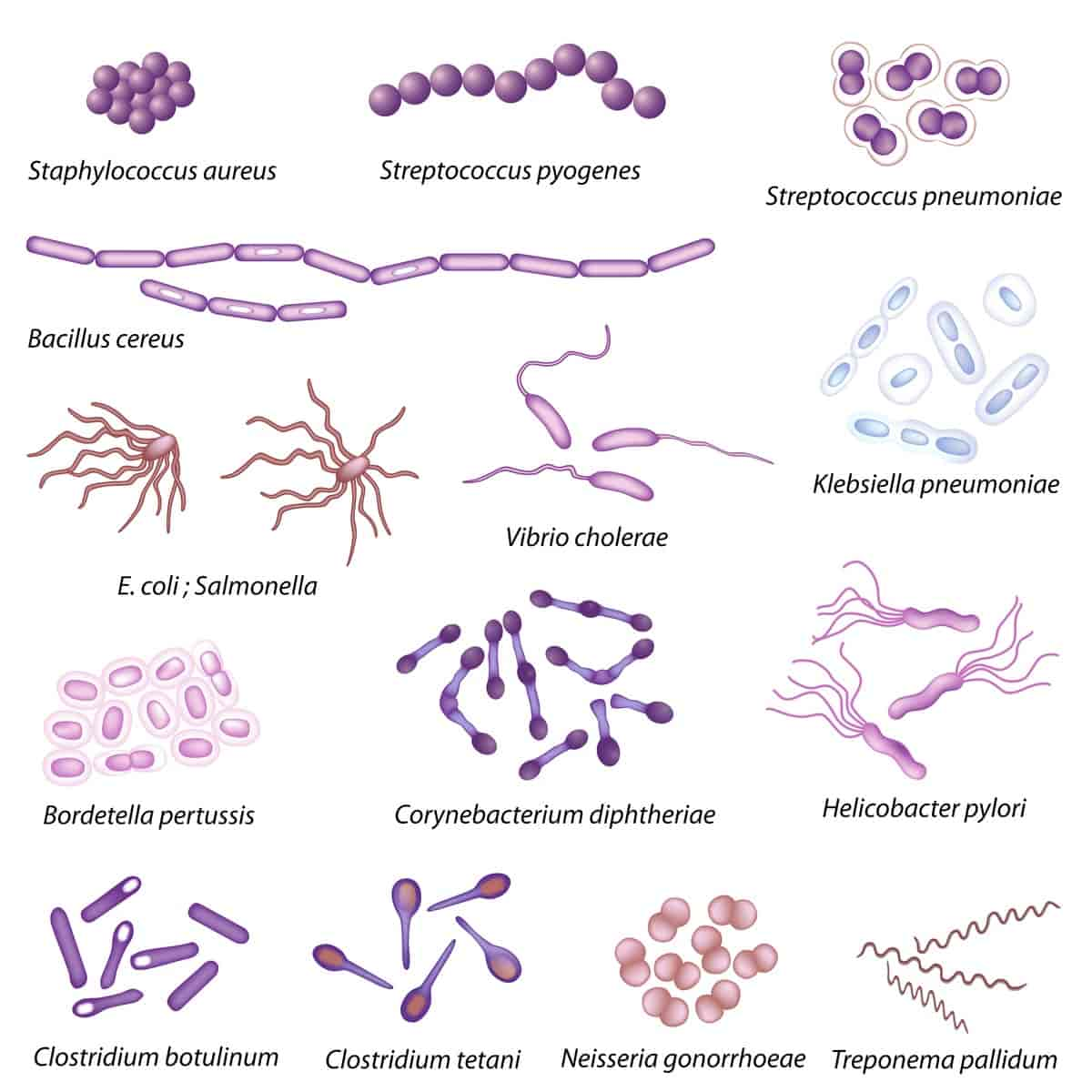
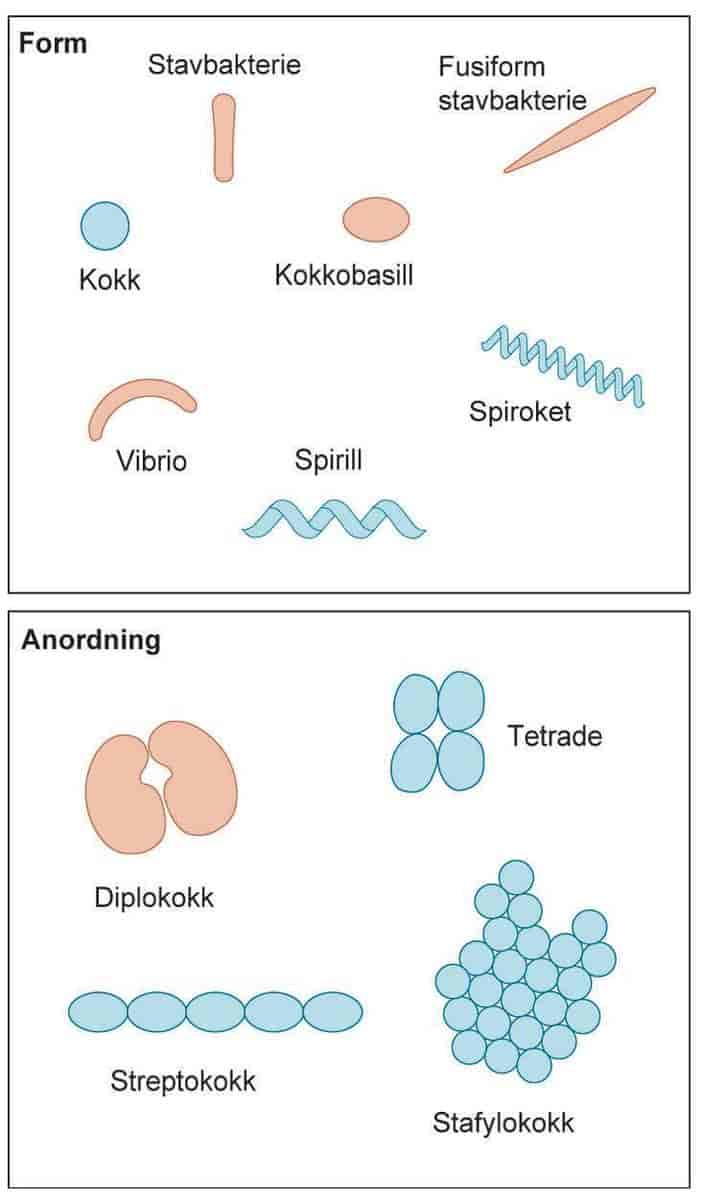
Bakterier er små encellete organismer som finnes overalt der det er mulighet for liv, som i jord, vann, luft, på og i andre organismer, i varme kilder og i forbindelse med radioaktivt avfall. Helt siden livet oppsto for omkring 3,8 milliarder år siden har det vært bakterier på Jorden. De er tilpasningsdyktige og har stor påvirkning på livet på Jorden.
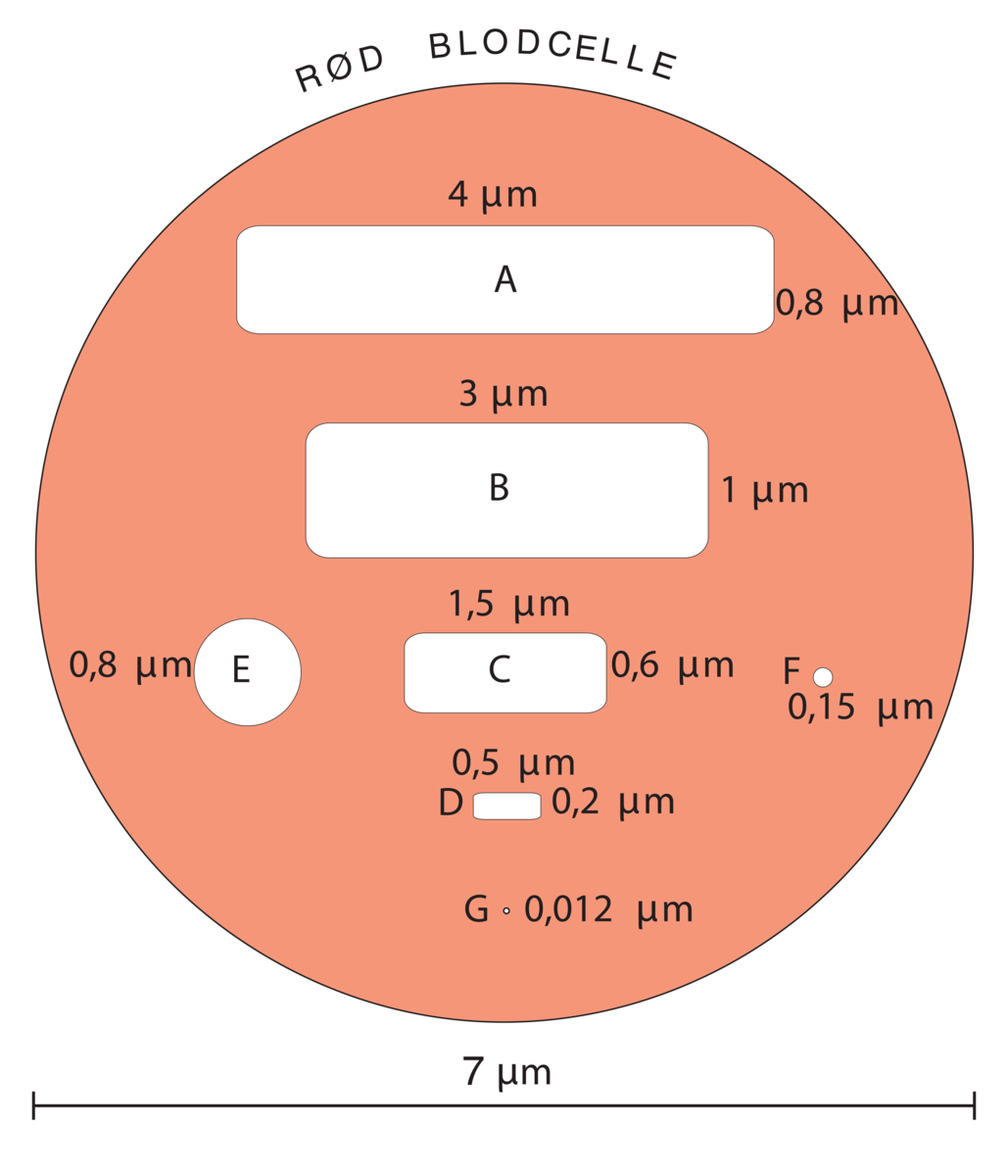
Størrelsen på bakterier varierer fra 0,2 til 10 mikrometer. De er så små at det er nødvendig med mikroskop for å se dem, og kalles derfor mikrober eller mikroorganismer. Encellete alger, protozoer og noen sopper er også mikroorganismer.
- Les mer om bakterieceller.
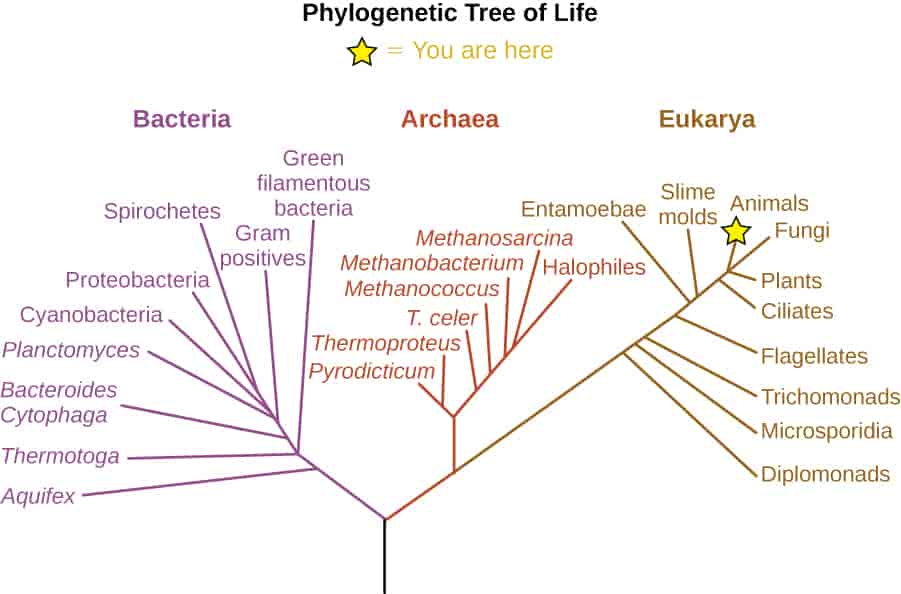
Bakteriene er prokaryote organismer som vil si at de ikke har avgrenset cellekjerne, men har arvematerialet sitt liggende løst inne i cellen. Dette i motsetning til eukaryote organismer, som planter, dyr og sopp, som har cellekjerne omgitt av en membran. Arker (Archaea) er også prokaryote.
Bakterier er ofte forbundet med sykdom og død, men de aller fleste bakterier er nyttige både direkte og indirekte. De påvirker oss på mange områder; helse, mat, landbruk, miljø, energi og industri.
Her er en liste over noen av de mest kjente bakteriene som forårsaker sykdom, ofte kalt sykdomsfremkallende bakterier:
| Organisme | Sykdom |
|---|---|
| Treponma pallidum | syfilis |
| Neisseria gonorrhea | gonoré |
| Salmonella typhosa | tyfoid feber |
| Vibrio cholera | kolera |
| Yersinia pestis | byllepest |
| Staphylococcus aureus | matforgifning, toksisk sjokk |
| Clostridium tetani | stivkrampe |
| Clostridim botulinum | botulisme |
| Legionella pneumonia | legionærsyke |
| Salmonella arter | matforgiftning |
| Clostridium perfringens | matforgiftning |
| Streptococcus arter | halsesyke, skarlagensfeber |
| Chlamydia trachomatis | klamydia, trakom |
Her er noen eksempler på bakterier som brukes i produksjon av medisiner, kjemiske forbindelser og næringsmidler:
| Organisme | Bruk i produksjon av |
|---|---|
| Streptococcus lactis | surmelk |
| Leuconstoc bulgaricus | yoghurt |
| Propionibacterium | sveitserost |
| Escherichia coli | insulinproduksjon |
| Acetobacyet suboxidans | vineddik |
| Clostridium acetobutylicum | butanol, aceton |
| Rhizobium | ammoniakk |
| Pyrococcus | varmestabil DNA-polymerase |
| Pseudomonas | bioremediering |


Kommentarer (3)
skrev Tor Stian Holte
svarte Kjell-Olav Hovde
skrev Lars Nygaard
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.