



Erter har spilt en sentral rolle i vår forståelse av genetikk. Den moderne arvelæren startet i 1866 da munken Gregor Mendel publiserte sine krysningsforsøk. Fra sine førsøk utledet Mendel egne lover om arv.
Valget av hageerter (Pisum sativum) som forsøksmateriale var meget gunstig, på grunn av en rekke egenskaper hageertene har.
- Erter er ettårige, og med lukkete og tokjønnete blomster er de selvpollinerende og selvfertile.
- Erter kan i tillegg krysspollineres kunstig ved å overføre pollen fra en plante til arrene i blomstene på en annen plante hvor pollenbærerne først er fjernet.
Dette gjorde det lett å kontrollere hvem som var foreldre og avkom for plantene som ble undersøkt.
- Ertene har en rekke egenskaper som er enten-eller, og dermed lettere å telle enn kontinuerlig varierende trekk.
- Det fantes mange konstante varieteter av erter.
Dette gjorde det lettere å måle og telle trekkene, og å være sikker på at resultatene skyldtes arv og krysninger, ikke for eksempel miljøet.
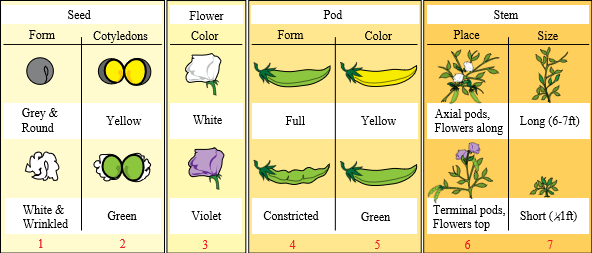
Egenskapene Mendel undersøkte var:
- belgenes form
- belgenes farge
- ertenes form
- ertenes farge
- blomstenes farge
- blomstenes plassering endestilt eller aksialt
- høyden på plantene
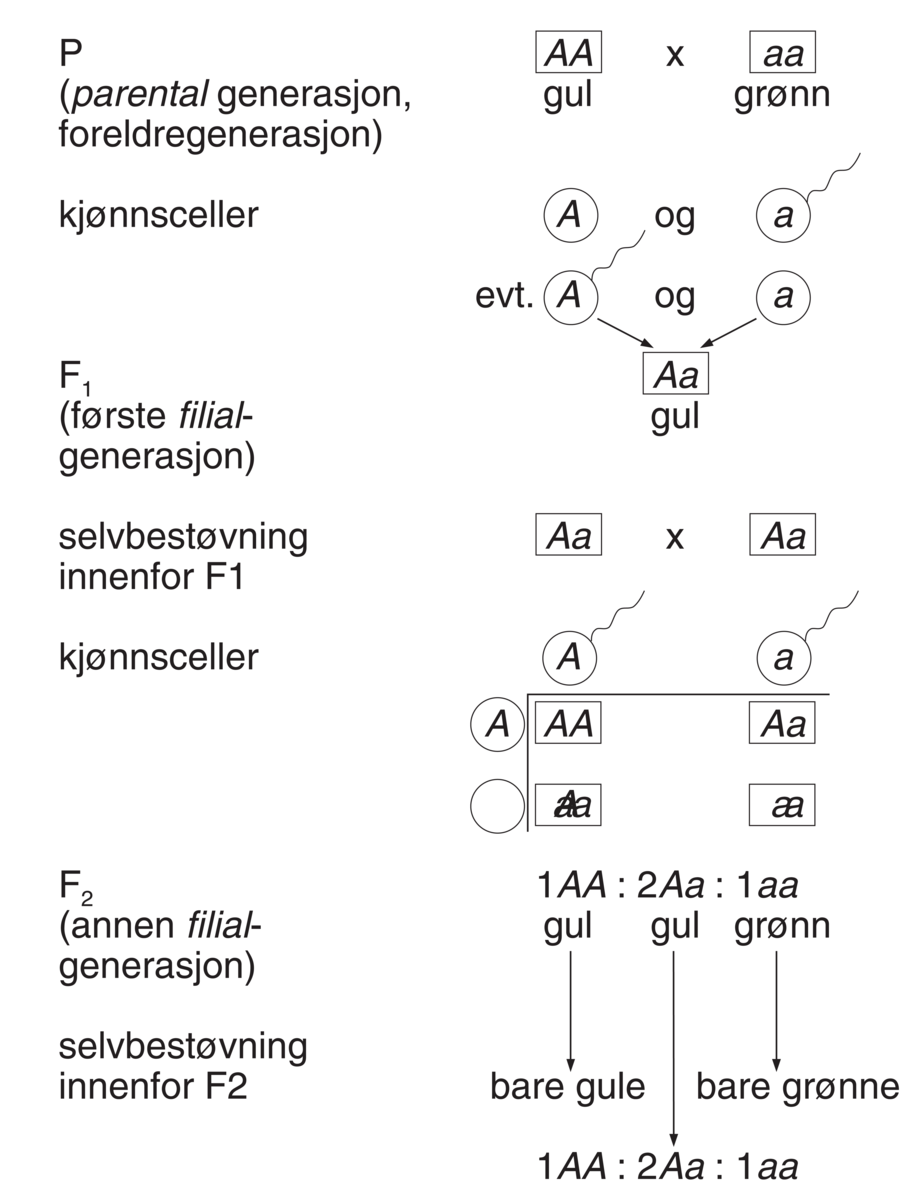
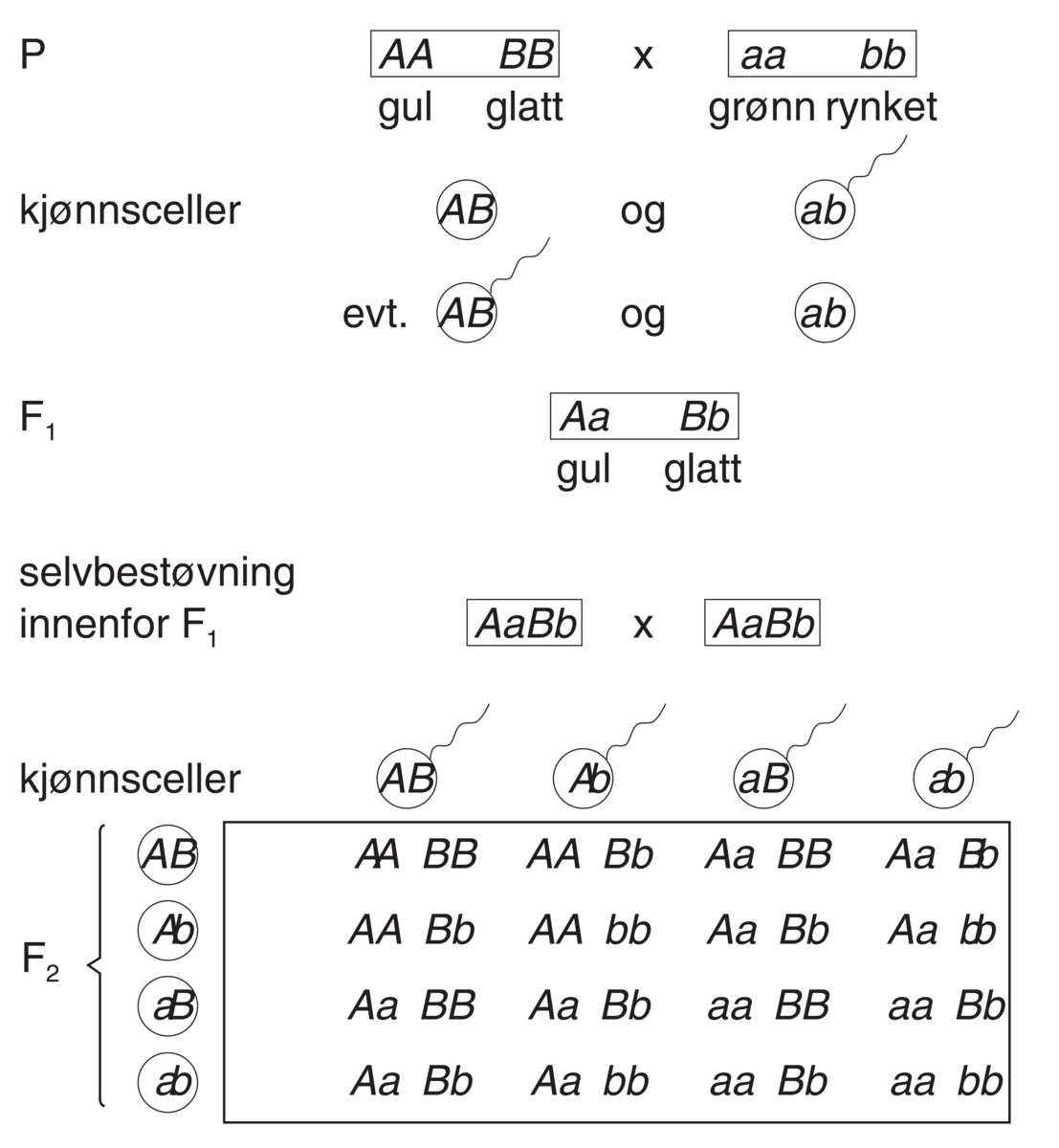

Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.