
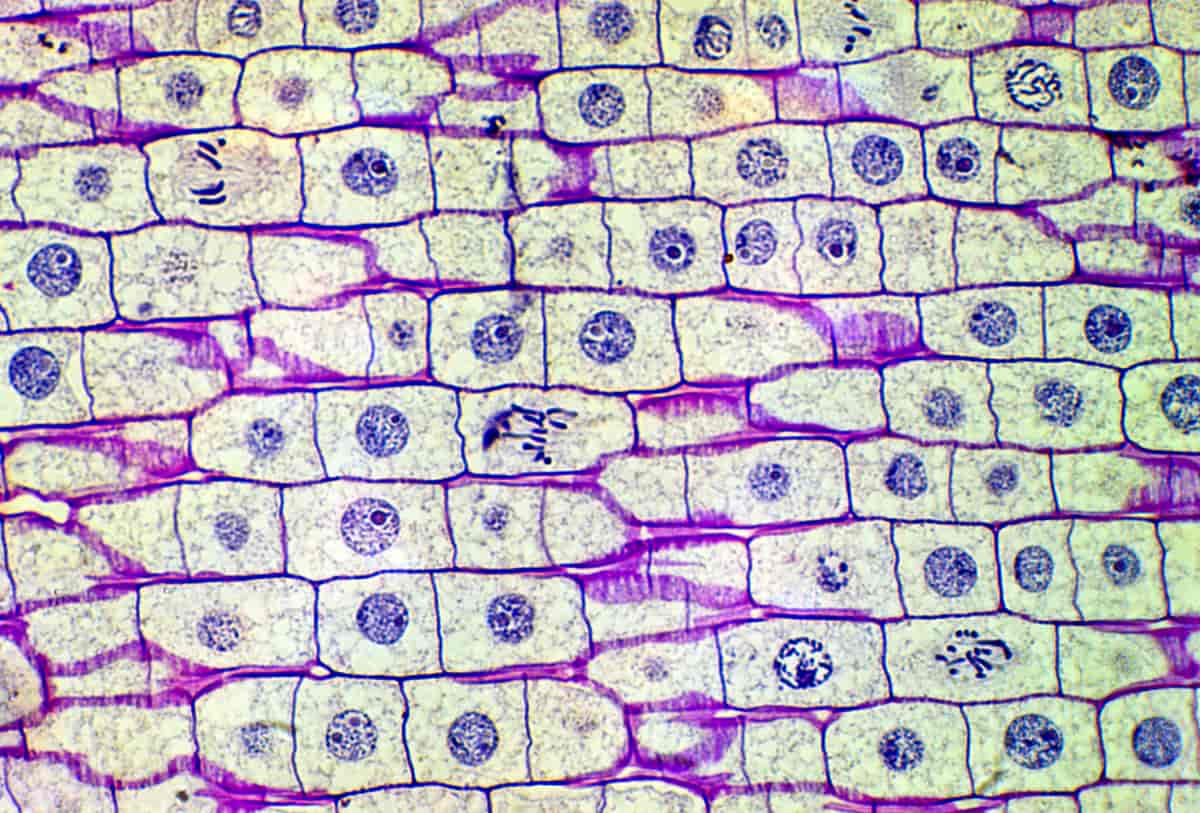
Celledeling er den prosessen hvor en celle deler seg og blir til to nye celler. Den cellen som deler seg kalles ofte morcelle, og de to nye cellene kalles datterceller. Celledeling er svært viktig i vekst, formering, utvikling og vedlikehold av organismer.
Avhengig av celletype og organisme skjer celledeling på ulike måter. Flercellete organismer som planter, dyr og sopp er bygget opp av celler der DNA-et ligger beskyttet i en cellekjerne (eukaryote celler). Disse cellene kan dele seg på to ulike måter: mitose og meiose.
Mitose er celledelingen som fører til at en celle deler seg og danner to nye datterceller med helt likt DNA. Disse dattercellene har da samme antall kromosomer som morcellen. Det er denne typen celledeling som brukes for vekst, erstatning og vedlikehold av vev og organer hos flercellete organismer.
Meiose er prosessen der det dannes kjønnsceller, det vil si sædceller og eggceller. I meiosen går morcellene gjennom to delingsrunder, meiose I og meiose II, og resultatet er fire datterceller. Hver av dattercellene har halvparten så mange kromosomer som morcellen. Når en eggcelle og en sædcelle smelter sammen, inneholder den befruktede eggcellen, også kalt zygoten, et normalt antall kromosomer for arten. Hos mennesket er dette 46 kromosomer. Etter dette deler zygoten seg utallige ganger ved hjelp av mitose.
Hver celledeling er nøye kontrollert for å sikre at cellene deler seg på riktig tidspunkt og på riktig måte. Feil i celledelingen kan føre til sykdommer som kreft hvor cellene deler seg ukontrollert.
Fra en morcelle starter å forberede celledelingsprosessen til cellen er delt i to datterceller tar det cirka 24 timer. Hvert sekund skjer det om lag 3,8 millioner celledelinger i en voksen menneskekropp.
Prokaryote celler (celler uten cellekjerne, bakterier) deler seg ved hjelp av en prosess som kalles todeling eller binær fisjon.

Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.