Fotorespirasjon skjer i en C2-oksidativ fotosyntetisk karbonsyklus, en biokjemisk omsetningsvei og en del av assimilasjonen av karbondioksid i fotosyntesen, og som skjer via samvirke mellom celleorganellene kloroplaster, peroksisomer og mitokondrier. Karbon (C), nitrogen (N) og oksygen (O) som inngår i syklusen blir resirkulert. I tillegg dannes aminosyrene glysin og serin som kan brukes til andre formål i stoffskiftet i plantene.
C2-oksidativ fotosyntetisk karbonsyklus finnes hos alle organismer med oksygenproduserende fotosyntese, planter, grønnalger, rødalger, brunalger og blågrønnbakterier. Blågrønnbakterier har imidlertid ikke celleorganeller og hos disse er syklusen litt annerledes.
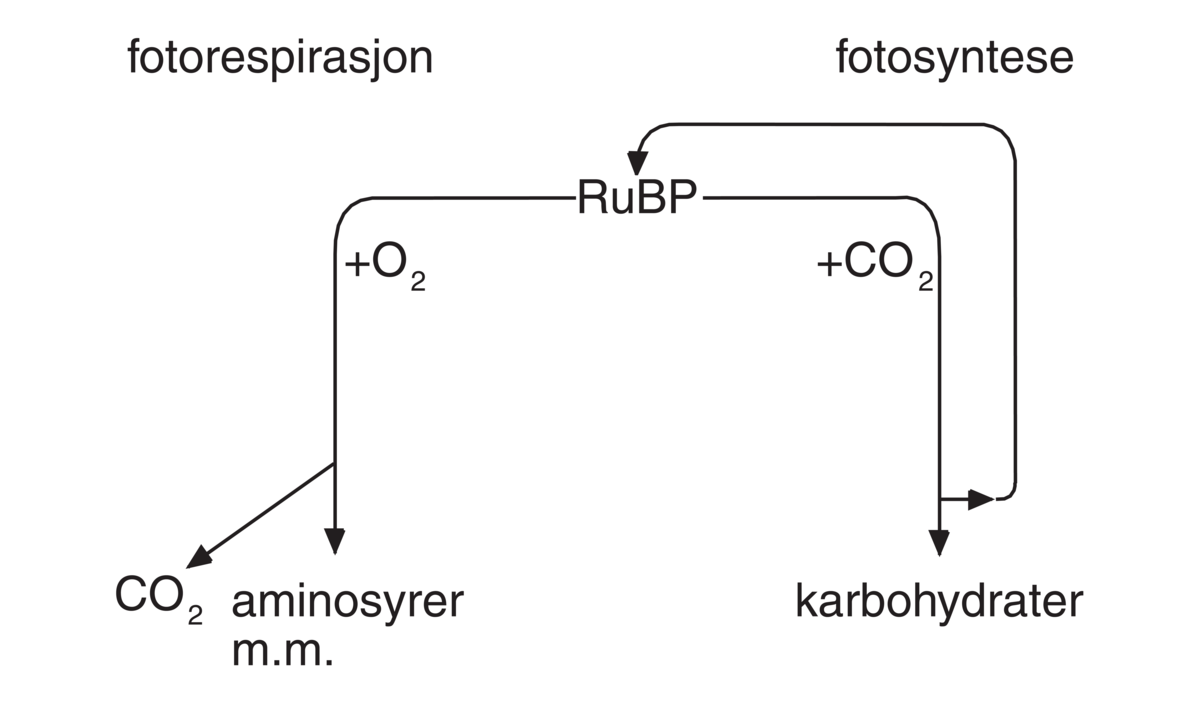
Substratet ribulosebisfosfat inneholder fem karbonatomer og når oksygen er substrat for rubisko blir produktet fosfoglycerat med tre karbonatomer, samt fosfoglykolat med to karbonatomer og denne er starten på C2 oksidativ fotosyntetisk karbonsyklus
ribulose-1,5-bisfosfat + O₂ → 2-fosfoglykolat + 3-fosfoglycerat
Fosfatgruppen på fosfoglykolat blir fjernet med hjelp av enzymet fosfatase:
2-fosfoglykolat + vann (H₂O) → glykolat + fosfat
Glykolat, anionet av glykolsyre (CH₂OHCOOH), overføres til en nærliggende organell, peroksysom.
I peroksysomene blir glykolat oksidert til glyoksylat katalysert av enzymet glykolat oksidase, et flavoprotein som inneholder FMN. I denne reaksjonen blir det laget hydrogenperoksid (H₂O₂) som et biprodukt, og dette er grunnen til at prosessen må skje i peroksisomene hvor hydrogenperoksid blir nedbrutt av oksygen og vann av enzymet katalase:
glykolat + O₂ → glyoksylat + H₂O₂
2H₂O₂ → O₂ + 2H₂O
Selv om hydrogenperoksid er en reaktiv oksygenforbindelse, fungerer den også som et signalstoff og redoksregulator.
Glyoksylat er en enkel ketosyre og den får hektet på en aminogruppe som kommer fra aminosyren glutamat katalysert av enzymet glutamat-glyoksylat-aminotransferase og produktene er aminosyren glycin (CH₂NH₂COOH) og en ketosyre, 2–oksoglutarat:
glyoksylat + glutamat ↔ glycin + 2-oksoglutarat
Glycin blir deretter transportert over til mitokondriene, hvor det er et stort enzymkompleks som i en oksidativ dekarboksylering omdanner to molekyler glycin til aminosyren serin, karbondioksid, ammonium (NH₄+) og reduksjonskraft (NADH).
Enzymkomplekset består av en glycin dekarboksylase og serin hydroksymetaltransferase og i reaksjonen inngår vitaminet folat i form av tetrahydrofolat (THF) . Folat er et vitamin som deltar i overføring av enkarbonforbindelser.
glycin + NAD+ + metylen-THF → NADH + NH₄+ + CO₂-THF-CH₂
glycin + CO₂-THF-CH₂ → serin + THF + CO₂
Karbondioksid går tilbake til kloroplastene og blir gjenbrukt som CO₂-kilde. Det samme gjør ammonium som er en viktig nitrogenkilde, men også kan virke giftig, gi ammoniumtoksisitet og blir raskt reassimilert.
Aminosyren serin fraktes tilbake til peroksisomene hvor aminogruppen blir overført til ketosyren 2-oksoglutarat katalysert av serin-2-oksoglutarat transaminase og gir hydroksypyruvat og aminosyren glutamat
serin + 2-oksoglutarat ↔ hydroksypyruvat + glutamat
Hydroksypuruvat blir redusert til glycerat katalysert av enzymet hydroksypyruvat reduktase
hydroksypyruvat + NADH + H+ → glycerat + NAD+
Glycerat blir fraktet tilbake til kloroplastene hvor den blir fosforylert katalysert av enzymet glycerat kinase
glycerat + ATP → 3-fosfoglycerat + ADP
3-fosfoglycerat kan deretter gå inn i calvinsyklusen.
Ammonium blir reassimilert i kloroplastene via enzymsystemet GS-GOGAT. Enzymet glutamin syntetase (GS) kobler sammen ammonium med aminosyren glutamat i en reaksjon som krever energi i form av ATP og lager aminosyren glutamin
glutamat + NH4+ + ATP → glutamin + ADP + fosfat (Pi)
Aminogruppen på glutamin blir overført til en ketosyre katalysert av enzymet ferredoksinavhengig gluatamat syntase (GOGAT). Ferredoksin deltar i transport av elektroner i elektrontransportkjeden i fotosyntesen:
glutamin + 2-oksoglutarat + 2 ferredoksinredusert + 2H+ → 2 glutamat + 2 ferredoksinoksidert
Glutamat kan deretter bli brukt til transaminering i peroksisomene.


Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.