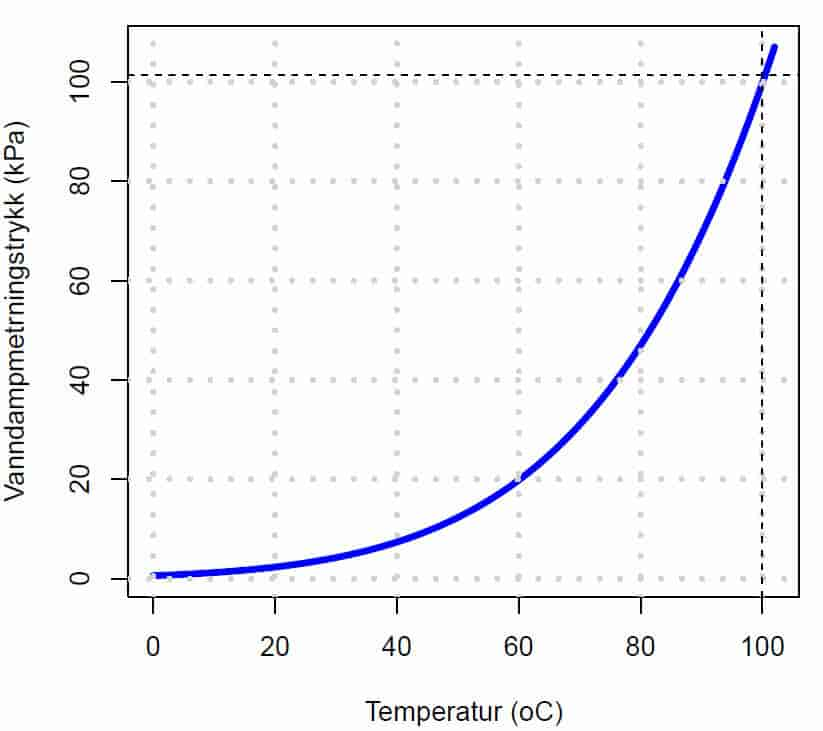
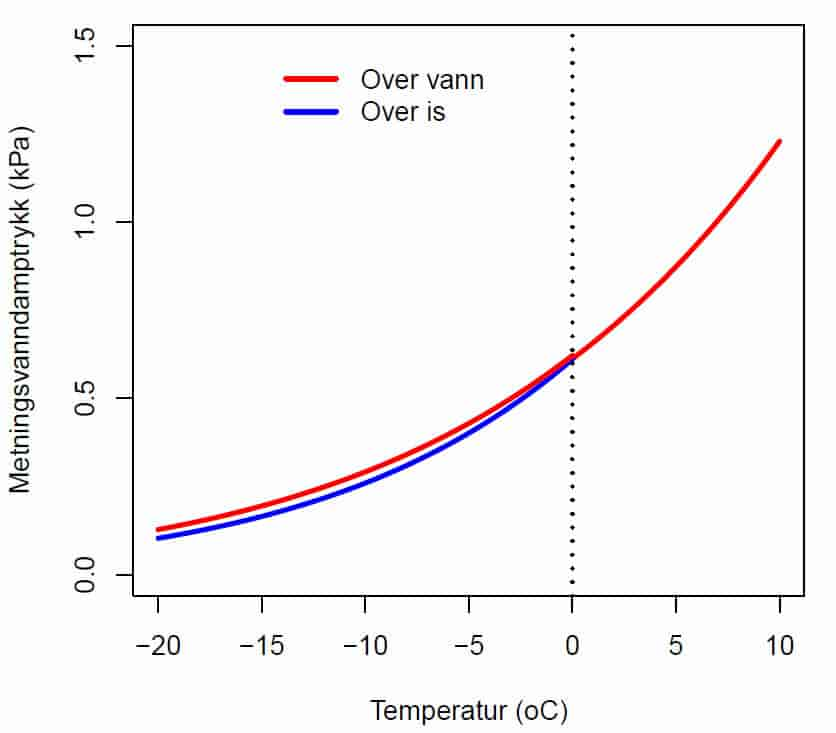
I et system med to reaksjonsrom ved samme temperatur atskilt av en halvgjennomtrengelig membran, så vil vannet bevege seg gjennom membranen til det reaksjonsrommet som har lavest vannpotensial. Rent vann har større evne til å utføre arbeid enn vann som inneholder oppløste stoffer.
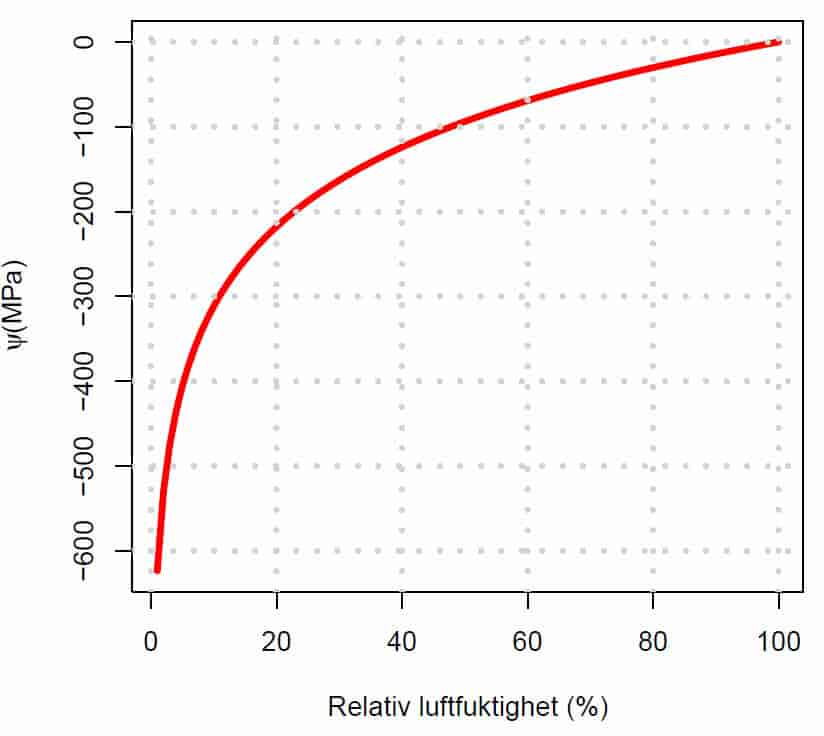
Størrelsen på vannpotensialet og de komponentene det består av varierer med art, i de forskjellige delene av planten, og under forskjellige vekstforhold. Vannpotensialet (ψ) i blader hos urteaktige planter er vanligvis i størrelsesorden -0,2 til -1,2 MPa, men i trær kan bladene ha vannpotensial opptil -2,5 MPa.
Siden en celle inneholder oppløse stoffer i en cytosol så vil det osmotiske potensialet (ψs) alltid være mindre enn null inne i en celle, for vanlige parenkymceller i størrelsesorden –0,5 MPa til –1,5 MPa. Derimot på utsiden av en celle, i apoplasten, hvor det er lite oppløste stoffer eller tilnærmet rent vann kan det osmotiske potensialet være fra –0,1 MPa til 0 MPa.
Sukkerbete, sukkerrør og vindruer hvor cellene kan inneholde mye oppløst sukker har osmotisk potensial (ψs ) inntil –2,5 MPa. Siden vanntransporten er passiv må vannpotensialet inne i rota være lavere enn vannpotensialet i jorda. I våt jord ved feltkapasitet er vannpotensialet i jorda –0,015 MPa –0,03 MPa.
Saltplanter (halofytter) har ofte tørketilpasset morfologi for eksempel sukkulens, for selv om de har rikelig tilgang på vann gjør saltet det vanskelig å få tak i vann. Saltplanter må ha lavere vannpotensial inne i røttene enn på utsiden i saltvann eller saltholdig jord. Saltplantene kan senke vannpotensielt ved å produsere osmolytter som senker det osmotiske potensialet og derved får et tilstrekkelig lavt vannpotensial i rotcellene slik at de kan ta opp vann fra sjøvann. Vannpotensial i sjøvann kan være i størrelsesorden –2,5 MPa.
Trykkpotensialet (ψp) i en plantecelle kan variere fra 0,1 MPa til 3 MPa. Når en plantecelle tørker ut minker turgortrykket i cellen og trykkpotensialet kan synke til 0 MPa. Hvis det skjer ytterliggere uttørking skjer det plasmolyse i cellene, plasmamembranen løsner fra celleveggen og henger bare fast i denne ved plasmodesmata, og cellen kan bli mekanisk deformert og dør. Inne i de døde cellene i vedvevet er trykkpotensialet negativt når det er transpirasjon gjennom bladene.

Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.