Mutasjoner er endringer i arvematerialet til en organisme. Slike mutasjoner skjer i alle organismer (dyr, planter, sopp, bakterier og virus) og er en naturlig prosess. Veldig viktig er også at endringene er tilfeldige. Ingen organisme kan fremskaffe mutasjoner for å tilpasse seg et miljø, men de mutasjonene som tilfeldigvis oppstår i gener som er viktige for organismen kan gjøre at den blir bedre tilpasset. Hos virus kan det for eksempel bety å kunne spre seg raskere. Dersom mutasjonen tilfeldigvis skjer i et gen som påvirker denne egenskapen vil viruset endre seg på en måte som får konsekvenser for verten, for eksempel mennesker. Veldig mange mutasjoner vil ikke ha noen påvirkning på viruset i det hele tatt fordi de skjer på områder i arvematerialet som ikke er viktige.
Det er to hovedårsaker til at mutasjoner oppstår: 1) Feil i cellenes egne funksjoner og 2) ytre påvirkning (for eksempel skadelig stråling og kjemikalier). Virus muterer altså ikke med hensikt for å bli mer dødelig eller spre seg mer.
Mutasjoner skjer veldig hyppig i virus. Ingen andre organismer muterer like mye som virus.
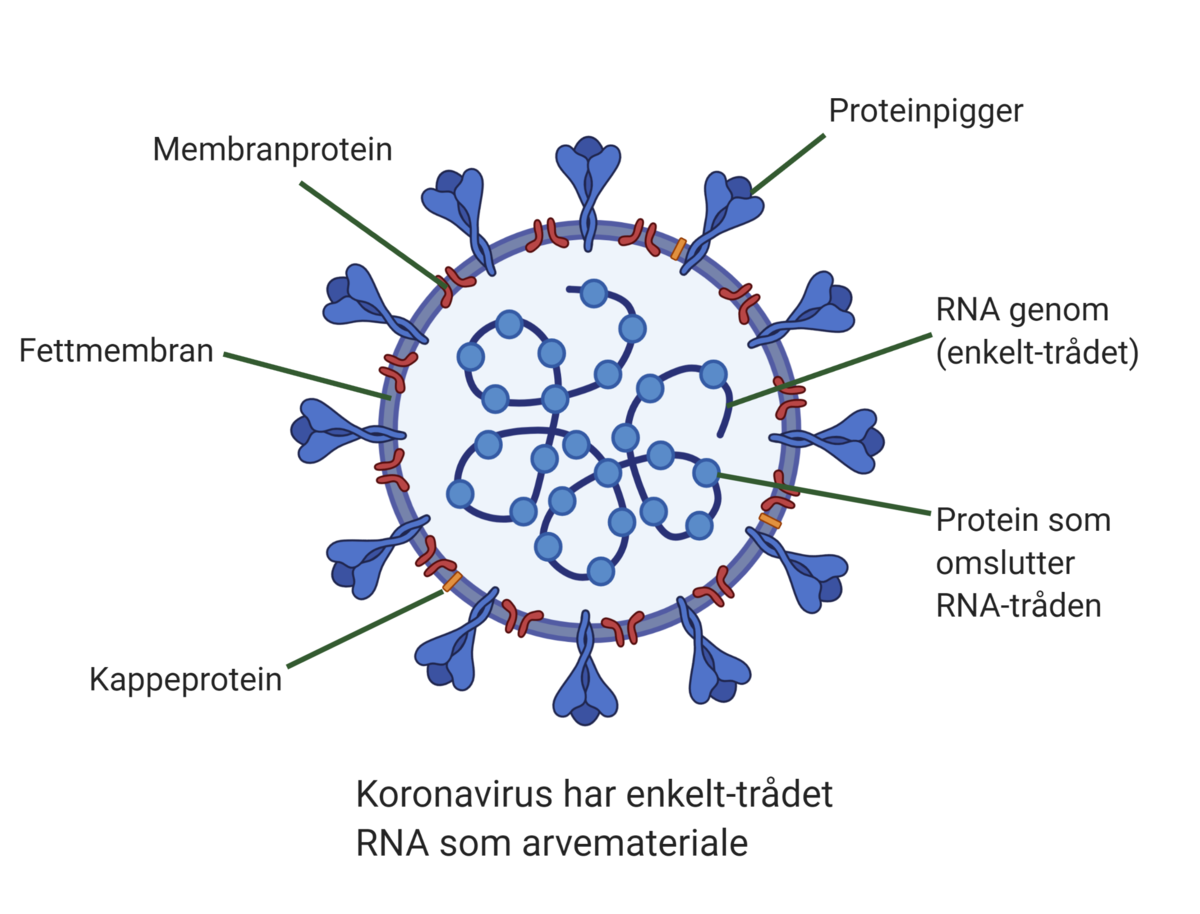
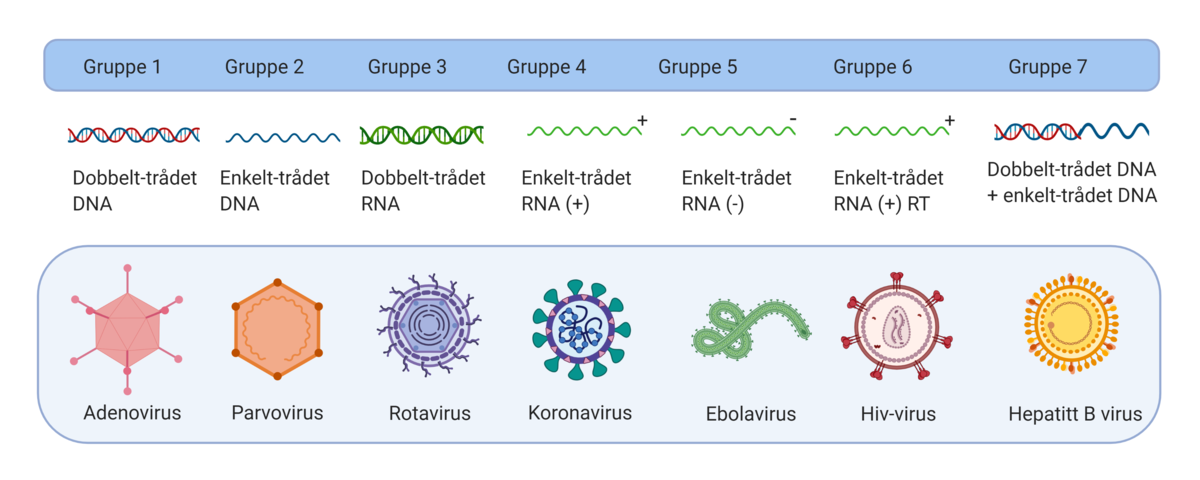
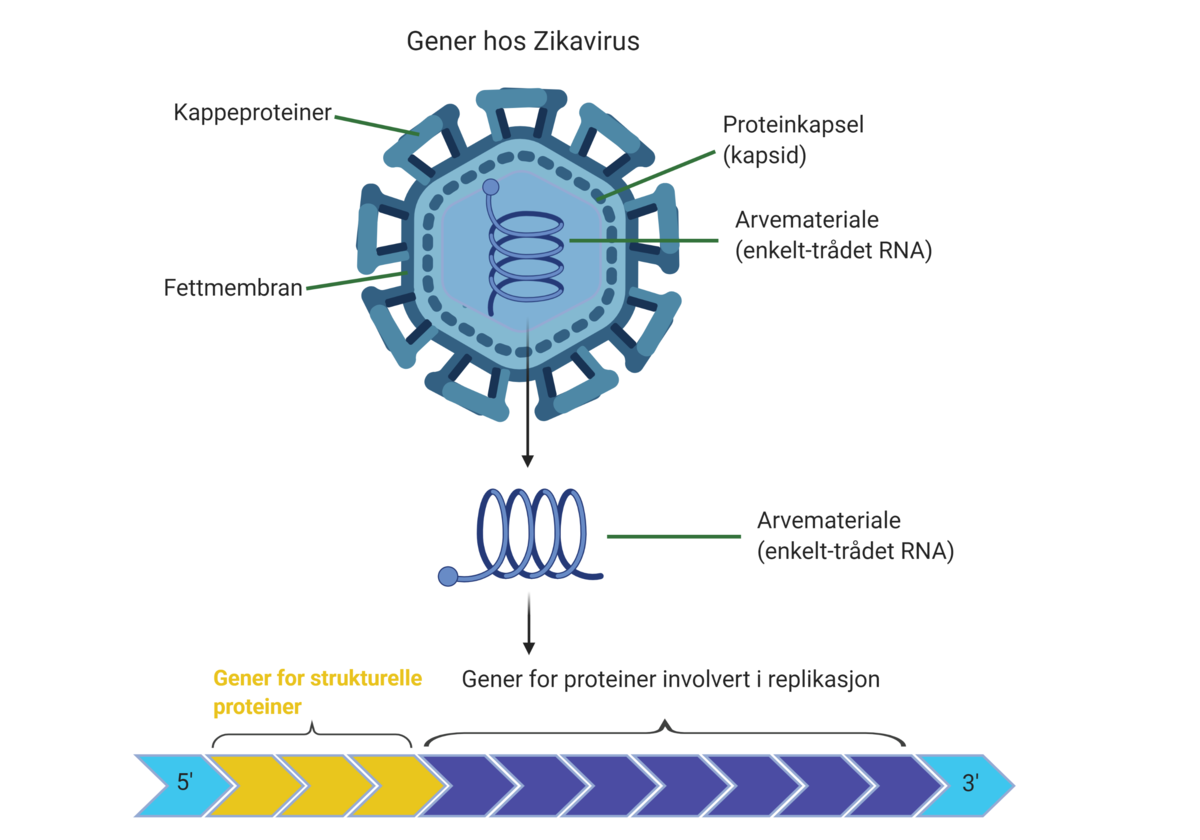
RNA-virus muterer oftere enn DNA-virus fordi RNA er et mindre stabilt molekyl enn DNA. RNA-virus har ikke de verktøyene som trengs for å reparere feil når arvematerialet kopier seg opp for å lage nye virus. Mutasjonsraten varierer mellom ulike virus, for eksempel ser det ut til at koronaviruset SARS-CoV-2 som forårsaker sykdommen covid-19 muterer langsommere enn influensaviruset selv om begge er RNA-virus (basert på kunnskap per april 2020). Det kan være en fordel ved utvikling av vaksine mot koronaviruset.
Ulike typer virus av samme art kan også utveksle gener seg imellom – for eksempel influensavirus. Dette øker muligheten for genetisk variasjon hos disse virusene.


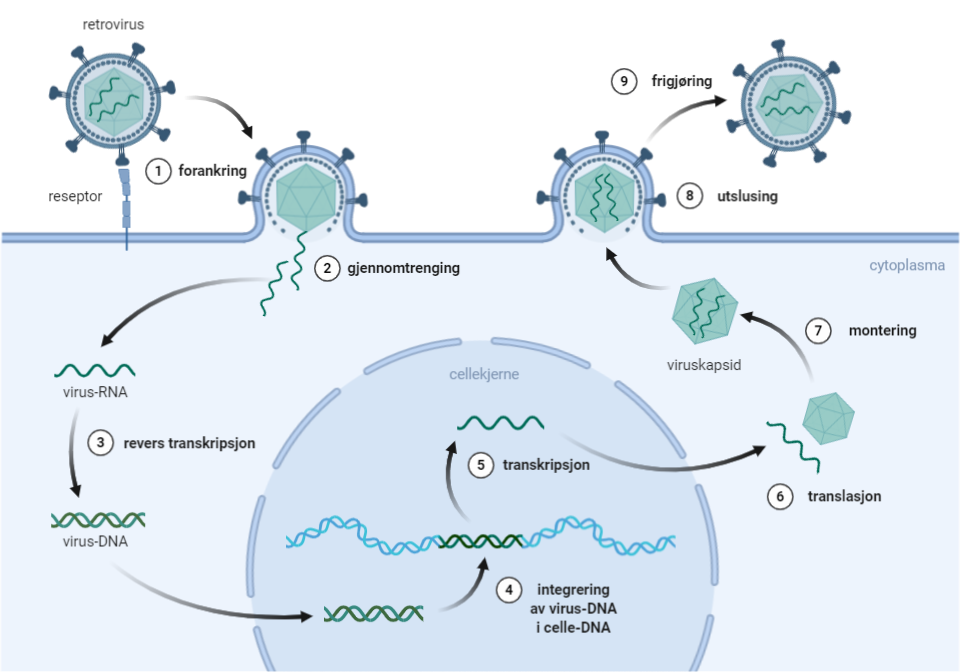
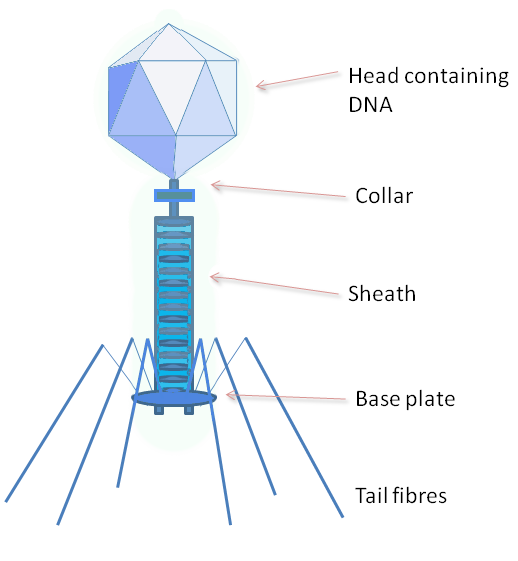
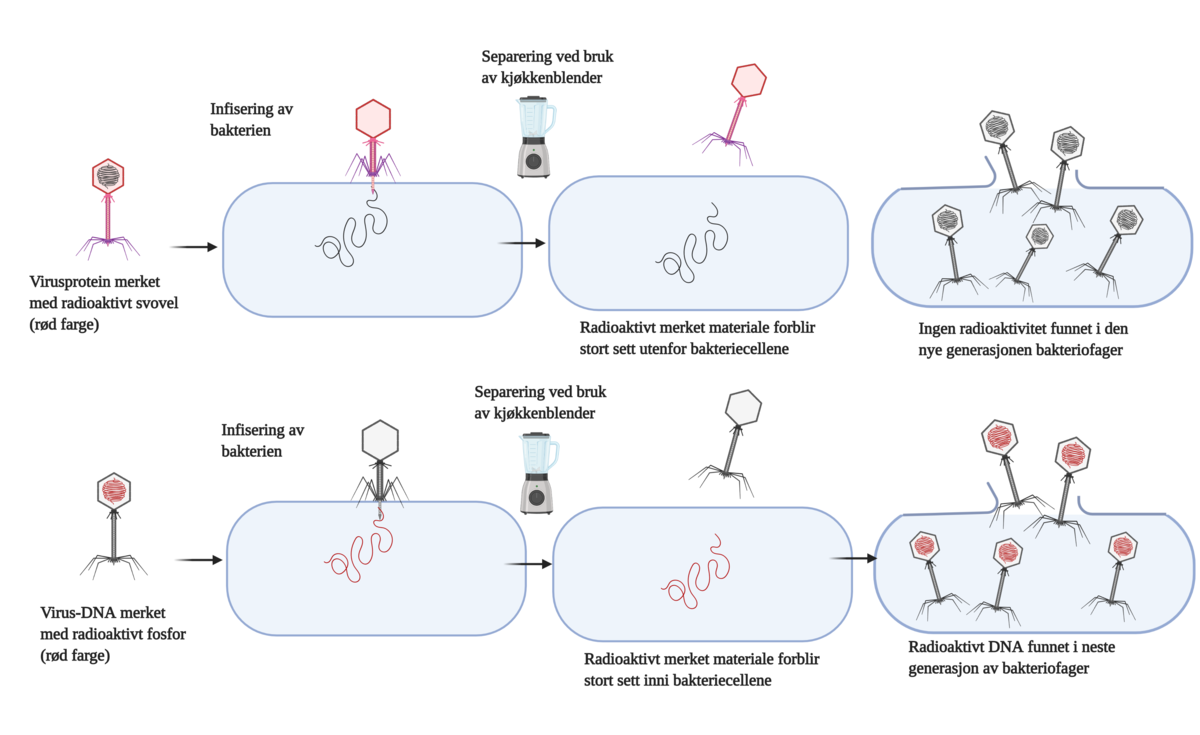

{kind=link}
Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.