
Nukleinsyrer er biokjemiske forbindelser (molekyler) i cellene som inneholder genetisk informasjon.
Faktaboks
- Uttale
- nukleˈinsyrer
- Etymologi
- til nukleus
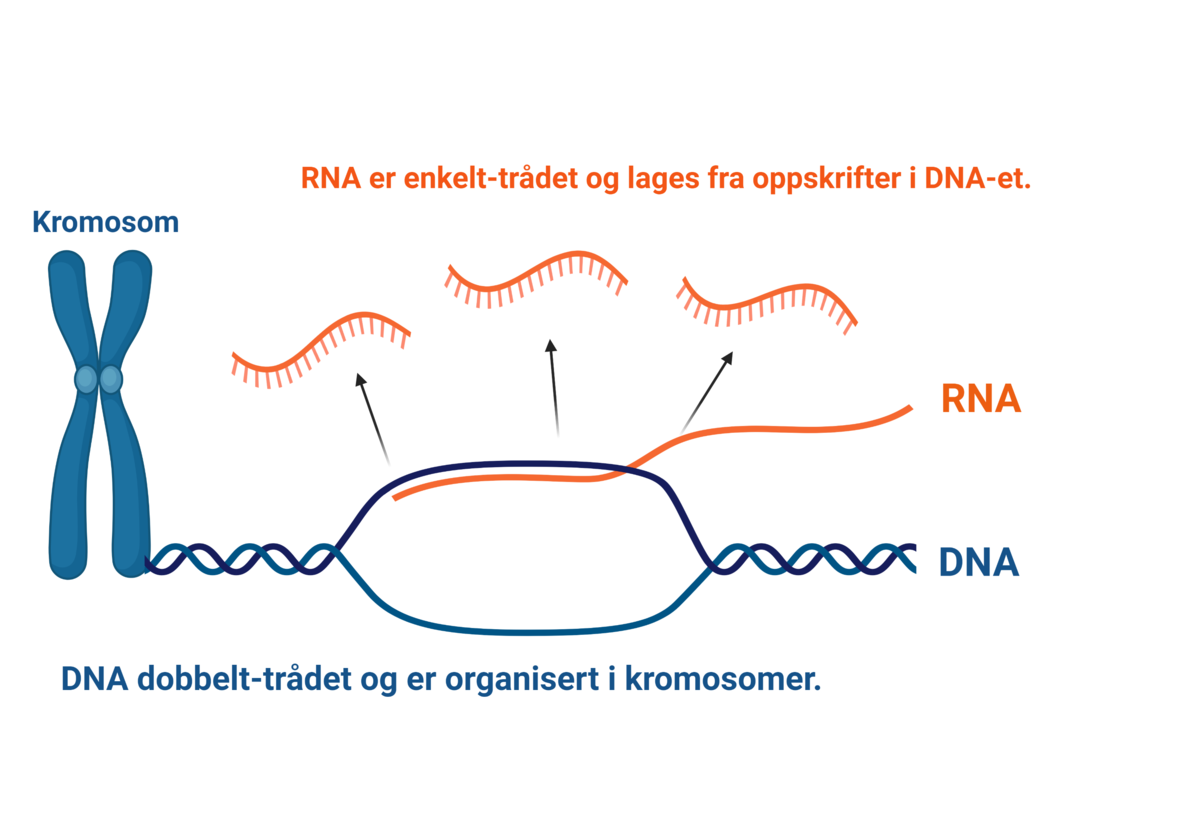
Nukleinsyrene er delt inn i de to hovedgruppene DNA og RNA.
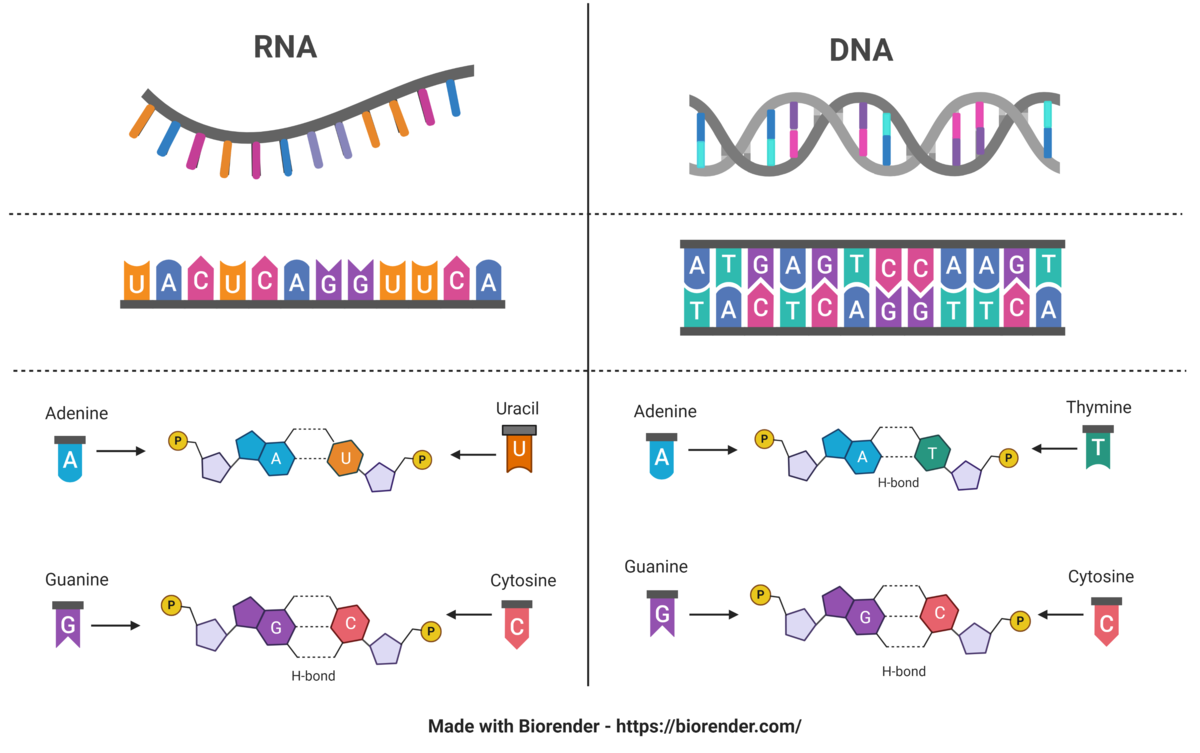
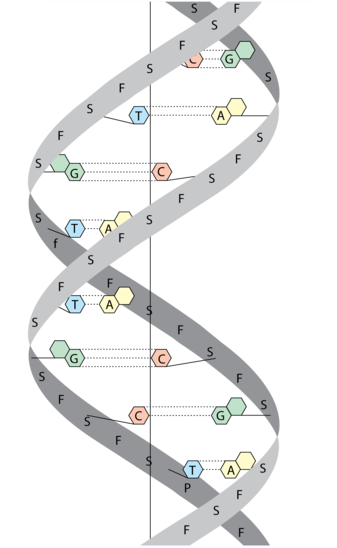
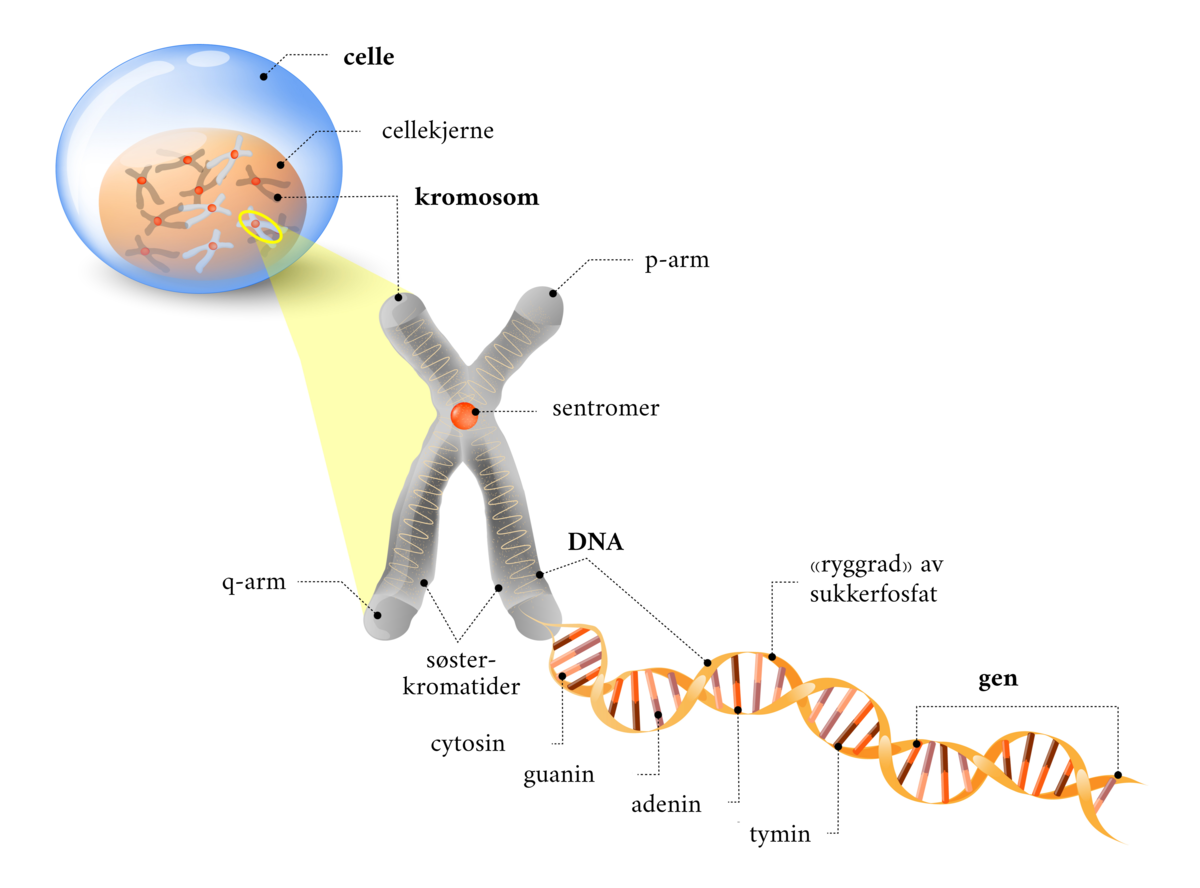
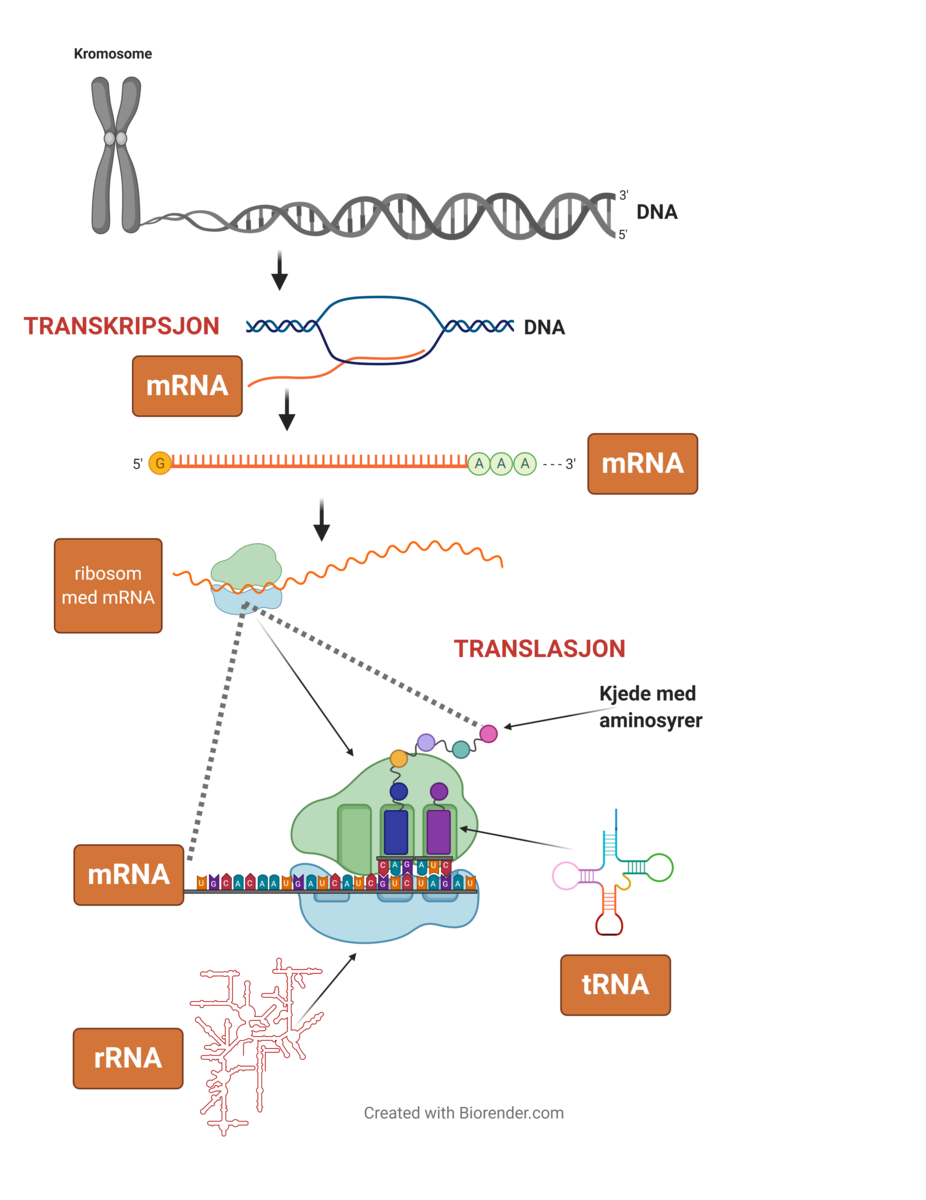
DNA er en forkortelse for det engelske navnet deoxyribonucleic acid (deoksyribonukleinsyre på norsk). DNA utgjør arvestoffet, det vil si alle genene, i en celle. DNA finnes hovedsakelig i cellekjernen i eukaryote celler, men også i mitokondrier, og i kloroplaster i planteceller. I bakterieceller er DNA i cytoplasmaet, siden disse ikke har cellekjerne. DNA er bygget opp av to tråder av nukleotider, og er formet som en dobbeltheliks.
RNA er forkortelse for det engelske navnet ribonucleic acid (ribonukleinsyre på norsk). RNA er molekyler som er bærere av genetisk informasjon i produksjon av proteiner og er viktig for regulering av hvilke gener som slås av og på (genregulering). RNA finnes hovedsakelig i cellenes cytoplasma. Det finnes flere typer RNA, blant annet tRNA, rRNA og mRNA som er viktige i proteinsyntesen. RNA består kun av én tråd av nukleotider.

Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.