
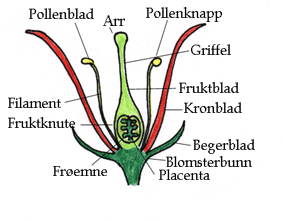
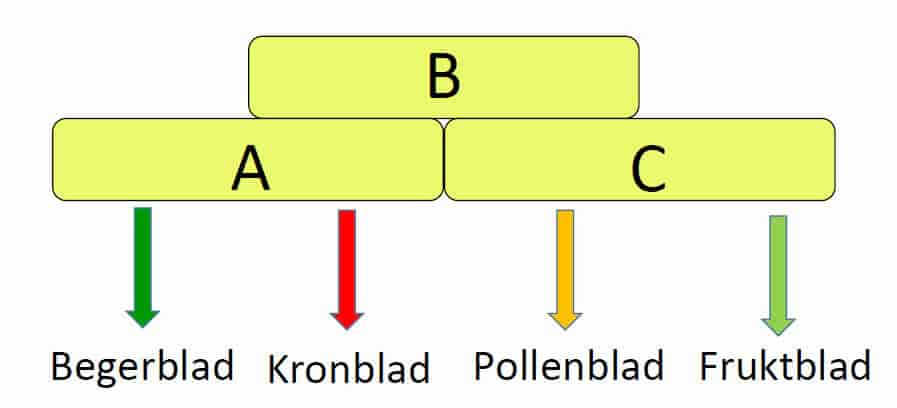
ABC-modellen består av tre klasser med gener A, B og C som koder for organidentiteten til de fire forskjellige bladene i en blomst. Klasse A koder for begerblad, klasse A + B er nødvendig for å lage kronblad, Klasse B + C gir pollenblad og klasse C gir fruktblad. Hos vårskrinneblom og løvemunn kjenner man til hvilke typer gener som inngår i A, B og C, hvorav flere er MADS-boksgener som koder transkripsjonsfaktorer. Modellen er nå utvidet til et kvartettmodell ABCE, hvor klasse E er gener som koder for blomstermeristem identitet.
Allerede Johann Wolfgang von Goethe i Die Metamorphose der Planzen var inne på idéen at de forskjellige delene av blomsten egentlig er forskjellig type blader.



Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.