




Blomsterplantene, eller de dekkfrøete plantene, er den mest artsrike og økologisk varierte av alle plantegrupper. De dominerer mesteparten av landjorda. Om lag 290 000 arter av blomsterplanter er kjent, men nye, tidligere ubeskrevne arter oppdages jevnlig, spesielt i tropiske strøk.
Faktaboks
- Også kjent som
- , Magnoliophytina dekkfrøete planter
- Vitenskapelig navn
- Magnoliophyta
- Beskrevet av
- Cronquist, Takht. og Walter Max Zimmerman
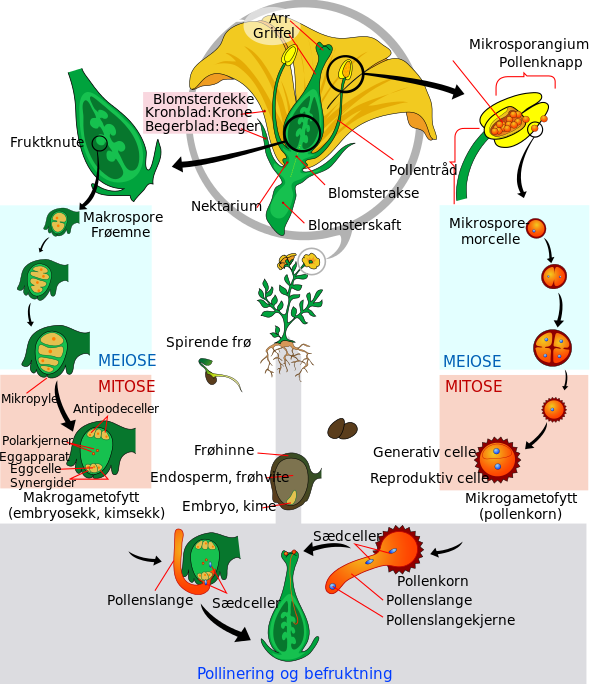
Blomsterplantene er frøplanter, som betyr at de formerer seg ved hjelp av frø. Blomsterplantene kalles også de dekkfrøete plantene fordi frøemnet utvikles i en slags beholder. Dette skiller blomsterplantene fra de nakenfrøete plantene, som også er frøplanter, men hos de nakenfrøete sitter frøemnene ganske åpent på et bladliknende organ. Blomsterplantene skiller seg også ved at de har ekte blomster og ved at de har såkalt dobbelt befruktning. De har i tillegg noen anatomiske spesialiseringer som forklares i egne avsnitt nedenfor.
Den viktigste evolusjonære nyvinningen er imidlertid den insektpollinerte blomsten. Med iøynefallende blomster og ulike belønninger for besøkende insekter har blomsterplantene evolvert sammen med insektene til stor diversitet.


Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.