
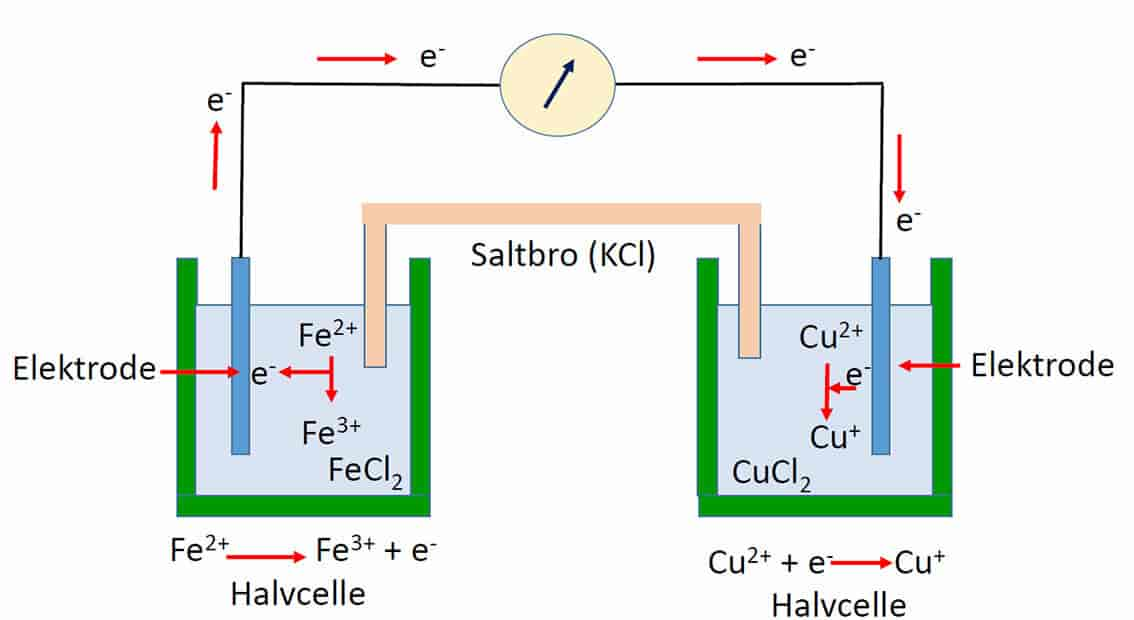
Biologiske redoksreaksjoner er redoksreaksjoner (egentlig reduksjons-oksidasjons-reaksjoner) som skjer i levende organismer. Redoksreaksjoner danner grunnlaget for livet på Jorden, siden det er en grunnleggende måte å lagre og å frigi energi på. Energi blir lagret når molekyler mottar elektroner i en reduksjon, og energi blir frigitt når molekyler avgir elektroner i en oksidasjon. En oksidasjon og reduksjon skjer alltid samtidig i reaksjonspar (redokspar). Når noe blir oksidert blir noe annet samtidig redusert, og omvendt.
Faktaboks
- Også kjent som
-
Biologiske reduksjons- og oksidasjonsreaksjoner, reduksjons-oksidasjonsreaksjoner
I celler skjer det for eksempel redoksreaksjoner som er koblet til protongradienter over mitokondrienes membraner, og det lages biokjemisk energi (ATP) og reduksjonskraft (evne til reduksjon). ATP brukes til å drive energikrevende prosesser i cellen.
| Lagre energi | Frigi energi |
|---|---|
| Reduksjon | Oksidasjon |
| Motta elektroner (+e-) | Avgi elektroner (- e-) |
| Motta protoner (+H+) | Avgi protoner (- H+) |
| Avgi oksygen (-O) | Motta oksygen (+ O) |
Et hydrogenatom (H) inneholder ett negativt ladet elektron (e-) og ett positivt ladet proton (H+) og selv om disse kan atskilles i enkeltreaksjoner vil de i sum følge hverandre og gi ladningsnøytralitet. De fleste av elektronene i atomer og molekyler er opptatt rundt atomkjernene, men elektroner som finnes i ytterste elektronskall eller ikke har noen fast plass, kan delta i elektronforflytningen i energireaksjoner.
Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.