Noen enzymer brukes hele tiden og er alltid tilstede i cellene, de er konstitutive. Enzymene som er nødvendige for å bryte ned glukose er for eksempel konstitutive i mange bakterier. At de er tilstede hele tiden, skyldes at glukose er vanlig i naturen og dermed lett tilgjengelig for bakterievekst.
Laktose, derimot, er et disakkarid som ikke er så vanlig, og de nødvendige enzymene for å utnytte dette sukkeret, blir bare produsert når laktose er tilgengelig. Et av disse enzymene er ß-galaktosidase som katalyserer omdannelsen av laktose til glukose og galaktose:
\[\ce{Laktose ->[ß-galaktosidase] glukose + galaktose}\\% chemistry \sffamily\bfseries\]
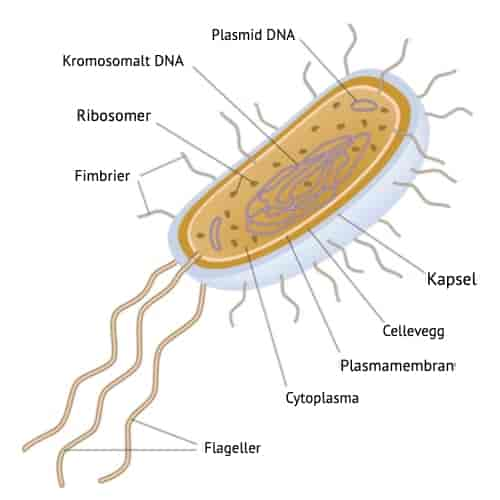
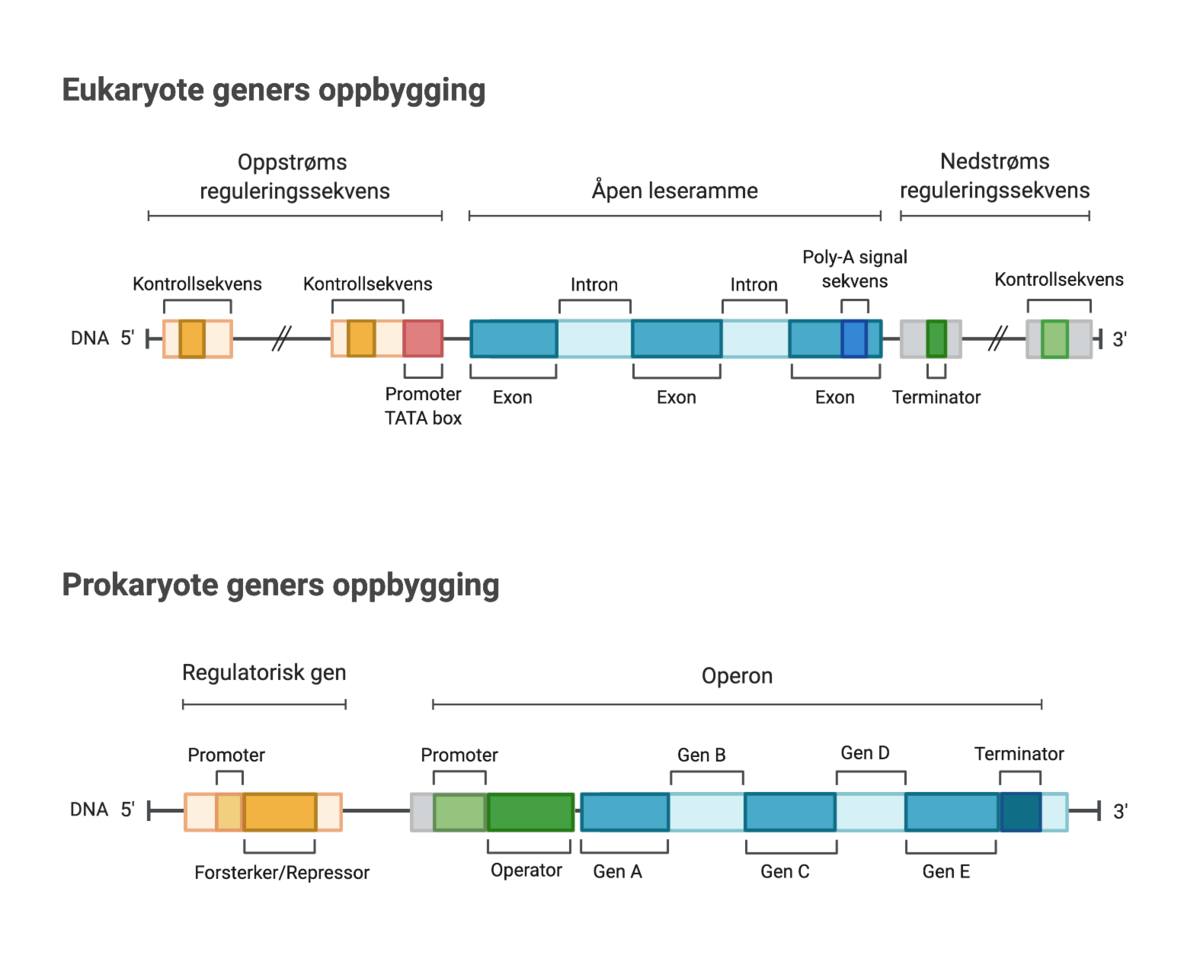
På kromosomet til E. coli ligger genet for ß-galaktosidase på en operon, lac-operon. Et operon er en gruppe gener som ligger sammen og er kontrollert av samme gen og transkriberes sammen. Genet for ß-galaktosidase sitter sammen med to andre, såkalte strukturgener som er nødvendige for vekst på laktose. Det ene er genet for en permease som transporterer laktose gjennom membranen og inn i cellen. Det andre koder for acetylase som er nødvendig for den videre omsetningen av galaktose. At de tre strukturgenene ligger samlet på en operon sikrer at de tre nødvendige proteinene er tilstede samtidig.
Like foran strukturgenene på lac-operon ligger genet for en regulator samt et område av DNA som kalles promoter og et som kalles operator. Promoter er stedet hvor RNA-polymerase starter transkripsjonen av genene. Regulator-genet koder for en repressor, et protein som binder seg til operator og hindrer RNA-polymerasen i å transkribere genene.
Under forhold hvor laktose ikke er tilstede eller ikke trengs, er repressoren bundet til operator og lac-operon transkriberes ikke.
Hvis cellen ikke har tilgang på glukose men på laktose, induseres lac-operon. Da binder laktosen seg til repressoren og fjerner den fra operator slik at transkripsjonen kan begynne.
Lac-operonet er i tillegg kontrollert av et annet protein, catabolitt aktivator protein (CAP). Transkripsjonen finner bare sted når CAP er bundet til promoteren, og CAP binder seg bare hvis nivået av cAMP (cyklisk AMP) i cellen er høyt. Det er høyt bare hvis det er lite glukose tilstede i cellen.
Dette betyr at lac-operon bare er aktiv når laktose er tilstede og glukose er nesten fraværende. Fordi transkripsjon av gener settes i gang ved induksjon er dette positiv kontroll.


Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.